Вирус оспы тип генома
Содержание статьи
еном вирусов оспы. Антигены вирусов оспы.
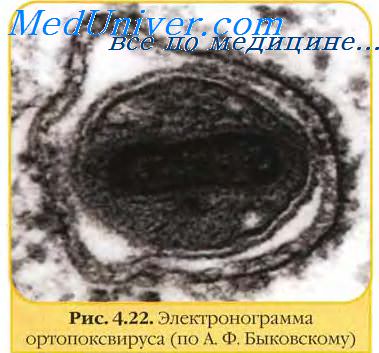
Геном вирусов оспы. Антигены вирусов оспы.Геном вирусов оспы представлен одной линейной молекулой двуцепочечной ДНК с ковалентно замкнутыми концами размером 130 тпн (парапоксвирусы) — 280 тпн (авипоксвирусы). На обоих концах генома имеются идентичные, но противоположно ориентированные тандемы повторяющихся нуклеотидных последовательностей. Геномы вирусов оспы способны кодировать около 200 белков, из которых не менее 100 входят в структуру вириона. Однако функциональные особенности определены лишь у небольшого количества вирусных белков. Наиболее важными из них являются ферменты, участвующие в синтезе вирусных нуклеиновых кислот и структурных компонентов вирионов. Например, синтез ДНК-полимеразы, ДНК-лигазы, РНК-полимеразы, энзимов, связанных с кэппированием и полиаденилированием мРНК и тимидинкиназы. Инфекционные вирусные частицы содержат системы транскрипции, которые могут выполнять in vitro синтез РНК, а также способны полиаденилировать, кэппировать и метилировать. В вирусных частицах содержится большое количество кодируемых вирусом энзимов и других биологически активных факторов. Некоторые гены вирусов оспы кодируют белки, которые секретируются инфицированными клетками и вызывают ответ организма на инфекцию, в том числе и формирование иммунитета. К таким вирокинам относятся гомологичный эпидермальный фактор роста, белок, снижающий активность комплемента, вирокины, обеспечивающие устойчивость к интерферону, и другие супрессоры иммунного ответа, подавляющие действие некоторых цитокинов организма хозяина. Вирусы оспы обычно характеризуются узким спектром хозяев. Они передаются чаще респираторным путем и реже через поврежденную кожу. Вирусы оспы овец, свиней, птиц и миксоматоза передаются также через укус членистоногими. Вирусы оспы устойчивы в окружающей среде и могут сохраняться годами в высохших струпьях кожи или других вируссодержащих материалах.
Большинство вирусов оспы хорошо размножаются в культуре клеток. Исключение составляют парапоксвирусы, вирус оспы свиней и вирус контагиозного моллюска. Однако они, так же как и ортопоксвирусы, легко образуют оспины на хориоал-лантоисной оболочке куриных эмбрионов. Вирусы оспы размножаются в цитоплазме, и, в отличие от других ДНК-вирусов, их размножение происходит независимо от ядра клетки, благодаря кодированию всех ферментов, необходимых для транскрипции и репликации вирусного генома. Некоторые из этих функций выполняются вирионами как таковыми. После слияния оболочки вириона с плазматической мембраной клетки или после эндоцитоза вирусная сердцевина освобождается в цитоплазму. Транскрипция вирусного генома характеризуется каскадностью, когда каждый временной класс генов («ранние», «промежуточные» и «поздние» гены) требует наличия специфических транскрипционных факторов, которые создаются предшествующим временным классом генов. Факторы, обеспечивающие транскрипцию промежуточных генов, кодируются ранними генами, тогда как факторы транскрипции поздних генов кодируются промежуточными генами. Транскрипция начинается вирионной транскриптазой и другими факторами, находящимися в сердцевине вириона, которые способны образовывать мРНК спустя минуты после инфицирования. Белки, образующиеся в результате трансляции этих мРНК, включая ДНК-полимеразу, тимидинкиназу и несколько других ферментов, необходимы для репликации вирусной ДНК. Репликация ДНК ВО связана с синтезом конкатемерных промежуточных структур, которые затем разрезаются с образованием единиц геномной длины. Детали этого процесса недостаточно изучены. С началом репликации ДНК происходит резкий сдвиг в генной экспрессии. Транскрипция «промежуточных» и «поздних» генов контролируется специфическими вирусными белками. Некоторые продукты транскрипции ранних генов образуются на поздней стадии инфекции, упаковываются в вири-оны и используются в следующем круге инфекции. Так как в состав вирусов оспы входит большое количество белков, не является неожиданным, что сборка вирионов есть комплексный процесс, который длится несколько часов и все еще целиком не выяснен. Образование вириона связано с вхождением ДНК внутрь незрелой сердцевинной структуры, которое затем завершается включением наружных покрывающих слоев. Репликация и сборка вирионов происходят в разных местах цитоплазмы в так называемых виропластах или вирусных фабриках. Вирионы выходят из клетки почкованием (оболочечные вирионы), или путем экзоцитоза, или при лизисе клеток (вирионы без оболочки). Большинство вирионов освобождаются при цитолизе и не имеют оболочки. Вирионы с оболочкой и без нее обладают инфекционностью, но первые, вероятно, играют более значительную роль в возникновении и распространении заболевания, а также в создании иммунитета. В очищенном вирусе осповакцины (ВОВ) выявлены белки с молекулярной массой 10-250 кД. Многие из них сосредоточены в сердцевине вириона. Два структурных гликопротеина располагаются между оболочкой и сердцевиной. В оболочке ВОВ содержится около 10 белков, из которых иммунологически наиболее активны крупномолекулярные белки с молекулярной массой 58—32 кД (VP4c, VP6a, VP6b и VP7a). Белок 32 кД определяет круг хозяев и важен для репликации вируса. В составе очищенного вируса оспы птиц обнаружено 29 полипептидов с молекулярной массой 14-138 кД. Наивысшей антигенной и иммуногенной активностью обладают полипептиды с молекулярной массой 35 и 37 кД. За индукцию вируснейтрализующих антител ВО ответственны антигены, расположенные на поверхности наружной оболочки вириона, и прежде всего белок 58 кД (VP4c), являющийся основным структурным компонентом трубочек (ворсинок). Антисыворотка к этому белку нейтрализовала инфекционность вируса и предотвращала образование синцития в культуре клеток. Этот белок ответственен за выработку иммунитета. Внеклеточные вирионы покрыты дополнительной наружной оболочкой, отсутствующей у внутриклеточных вирионов. Она играет важную роль в индукции синтеза ВН-антител. Инфекционность ВОВ и ВО крупного рогатого скота, имеющих наружную оболочку, нейтрализовалась антисывороткой к имеющему эту оболочку ВОВ, но не нейтрализовалась антисывороткой к ВОВ, лишенному наружной оболочки. За гемагглютинирующую активность ортопоксвирусов ответственны полипептид 85 кД и гликопротеид 41 кД. Во внеклеточном оболочечном ВОВ гемагглютинирующие свойства связаны с полипептидом 85 кД. Внутриклеточные вирионы (без дополнительной оболочки) практически не содержали этого полипептида. Неструктурный гемагглютинин формируется на цитоплазматических мембранах. С его образованием инфицированные клетки приобретают способность адсорбировать эритроциты. Вирусспецифические белки с молекулярной массой 32 и 37 кД, экспрессируемые на поверхности клеток, инфицированных ВОВ, делают их мишенями для специфических цитотоксических Т-лимфоцитов. МАТ-реактивные против каждого из пяти (54; 34; 32; 29 и 17—25 кД) белков наружного слоя поверхности вируса осповакцины нейтрализовали его инфекционность. В структуре полипептида 54 кД обнаружено два нейтрализующих эпитопа(Аи В). Анализ антигенных детерминант поверхностных полипептидов, проведенный с помощью МАТ, выявил в составе ортопоксвирусов, наряду с видоспецифическими, группоспецифические эпитопы. — Также рекомендуем «Оспа овец. Оспа птиц. Особенности оспы овец и птиц.» Оглавление темы «Частная вирусология.»: |
Источник
имический состав вирусов. Вирусные геномы. Виды вирусных геномов.
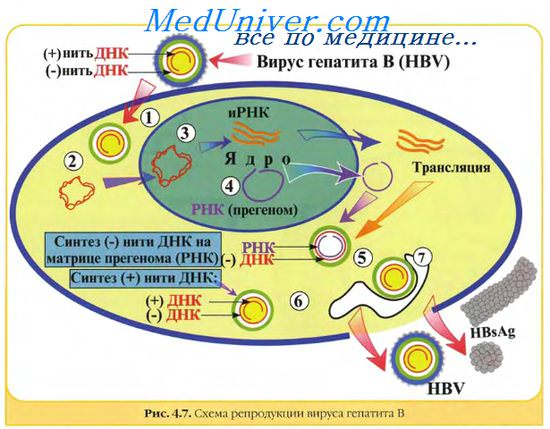
Химический состав вирусов. Вирусные геномы. Виды вирусных геномов.Химический состав вирусов отличается от других форм жизни необычайной простотой. Кроме геномной ДНК или РНК вирусы позвоночных содержат белки, масса которых составляет 57—90% массы вириона. Количество вирионных белков может колебаться в широких пределах в зависимости от сложности строения вируса. Среди белков, кодируемых вирусным геномом, различают структурные и неструктурные вирусспецифические белки. Первые входят в структуру вириона, вторые не входят. Структурными белками являются капсидные белки, белки оболочки и в некоторых случаях белки тегумента и ферменты. У многих вирусов некоторые структурные вирусные белки не являются первичными продуктами генов, а образуются в результате посттрансляционного расщепления полипептида-предшественника вирусспецифическими и клеточными протеазами. Неструктурными вирусными белками являются белки, кодируемые вирусным геномом, но не являющиеся составными частями вириона. Одни из них участвуют в репликации вирусных компонентов, другие — в сборке вирионов, третьи нарушают нормальный клеточный синтез. Они включают различные типы репликаз, связанные с репликацией ДНК, транскриптазы, транскрибирующие мРНК на геномной дцДНК или дцРНК или (-)оцРНК, вирусные протеазы, геликазы и лигазы. У ретро- и гепаднавирусов найдена обратная транскриптаза, транскрибирующая ДНК на матрице РНК. У ретровирусов найден уникальный фермент, участвующий в интеграции ДНК-продукта в хромосомальную ДНК клетки. Вирусы оспы, которые размножаются в цитоплазме и имеют много уникальных ферментов для процессинга РНК-транскриптов и репликации вирусной ДНК, мало зависят от метаболизма клетки. Вирусные белки имеют молекулярную массу 5-200 кД. Наиболее просто устроенные вирусы (вирусы-сателлиты, дефектные вирусы) кодируют синтез только одного белка, многие патогенные вирусы кодируют синтез 5—10 белков, крупные вирусы, такие как вирусы оспы, герпесвирусы, кодируют синтез до 200 белков. Хотя это немного по сравнению с клетками прои эукариотов (кодируют соответственно более 5000 и 100000 белков).
Вирусные геномы. Виды вирусных геномов.Все вирусные геномы являются гаплоидными, т.е. содержат одну копию каждого гена. Исключение составляют ретровирусы, которые обладают диплоидным геномом. Геномы ДНК-вирусов позвоночных представлены одной двуспиральной молекулой за исключением парво- и цирковирусов. Геномы полиома-, папиллома-, гепадна- и цирковирусов представлены кольцевой ДНК. ДНК гепаднавирусов частично двуспиральная, частично односпиральная. ДНК вирусов полиомы и папилломы является суперспиральной. Большинство линейных вирусных ДНК обладает способностью приобрести циркулярную конфигурацию, которая требуется для репликации по вращающемуся кольцевому механизму. Две цепи ДНК вируса оспы ковалентно связаны своими концами и при денатурации образуют большое одноцепочечное кольцо. У некоторых ДНК-вирусов (так же как у РНК-ретровирусов) имеются концевые повторяющиеся последовательности. Инвертированные концевые повторы обнаружены у адено- и парвовирусов. У адено-, гепадна- и парвовирусов, так же как у некоторых РНК-вирусов (пикорна- и калицивирусов), с 5′-концом генома ковалентно связан белок, играющий важную роль в его репликации. Все РНК-вирусы позвоночных за исключением рео- и бирнавирусов имеют одноцепочечные геномы. Геном некоторых РНК-вирусов состоит из нескольких (2-12) уникальных фрагментов, каждый из которых кодирует, как правило, один белок. РНК-вирусы с односпиральным геномом могут иметь различную полярность. Если они имеют ту же полярность, что и мРНК, то они могут прямо индуцировать синтез вирусного белка и считаются положительно (+) полярными. Если геномная нуклеотидная последовательность комплементарна мРНК, то они считаются отрицательно (—) полярными. К ним относятся: парамиксо-, рабдо-, фило-, ортомиксо-, арена- и буньявирусы. Все они имеют вирионную РНК-зависимую полимеразу (транскриптазу), которая в инфицированной клетке транскрибирует положительно-полярную РНК на матрице геномной вирусной РНК. У аренавирусов, по крайней мере, у одного рода буньявирусов, один из РНК-сегментов является двуполярным. Обычно у (+)полярных РНК-вирусов З’-конец имеет polyA-последовательность, а 5′-конец имеет кэп-структуру. Размер геномов РНК-вирусов (одноцепочечных 1,7—21 т.н.; двуцепочечных — 18—27 т.п.н.) значительно меньше размера генома многих ДНК-вирусов. Поэтому РНК-вирусы, как правило, кодируют меньше белков, чем ДНК-вирусы. Масса генома различных вирусов находится в пределах от 1 % (орто- и пара-миксовирусы) до 32% (парвовирусы) от массы вириона. Различные семейства вирусов позвоночных значительно различаются по структуре и функции генома. Основные типы вирусных геномов можно представить следующим образом: 5) частично двуцепочечной кольцевой незамкнутой молекулой ДНК (гепаднавирусы); Молекулярная масса ДНК различных вирусов позвоночных варьирует в широких пределах: от 0,7—1,5 МД у цирковирусов и парвовирусов, до 150—200 МД у вирусов оспы. Молекулярная масса генома у РНК вирусов колеблется менее значительно — от 2,0 до 20,0 МД. — Также рекомендуем «Классификация вирусов. Современная номенклатура в вирусологии.» Оглавление темы «История вирусологии. Основы вирусологии.»: |
Источник
Геномы вируса оспы из Древней Руси и Западной Европы удлинили генетическую историю заболевания на 1000 лет
Изображение:
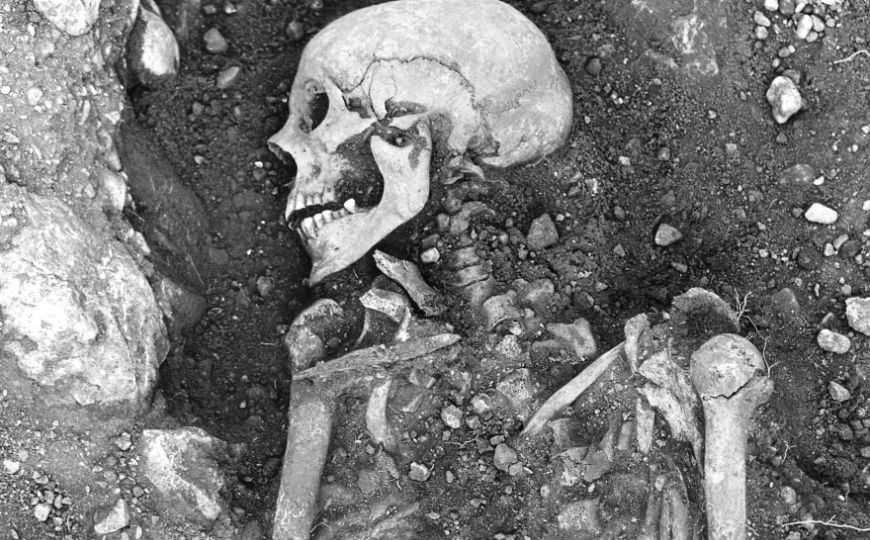
Скелет с острова Эланд (Швеция). Викинг, живший 1200 лет назад, был инфицирован вирусом оспы.
Credit: The Swedish National Heritage Board
Исследователи из разных стран проанализировали древние геномы возбудителя натуральной оспы — одной из самых опасных вирусных инфекций, только в ХХ векеубившей сотни миллионов людей. В конце ХХ века оспа была побеждена благодаря вакцинации; последний случай был зафиксирован в 1977 году.
Описания заразной болезни, похожей на оспу, были сделаны еще до нашей эры; есть находки костей с признаками оспы из Египта времен фараонов. Однако поскольку оспа редко поражает костную ткань, трудно получить прямые палеоанатомические свидетельства. До сих пор были секвенированы геномы штаммов вируса оспы возрастом до 360 лет, и последний их общий предок мог существовать в XVI–XVII вв. Таким образом, не существовало и прямых генетических подтверждений, что древние эпидемии, описанные в источниках, были именно оспой.
Международная группа под руководством вирусолога Терри Джонса из Института вирусологии клинического центра Шарите (Берлин) и палеогенетика Мартина Сикоры из университета Копенгагена вместе с Барбарой Мюлеман из Шарите провела поиск древних геномов вируса оспы. В исследовании приняли участие российские ученые — сотрудники НИИ и музея антропологии МГУ, истфака МГУ и Музея антропологии и этнографии им. Петра Великого (Санкт-Петербург).
Ученые выполнили высокопроизводительное секвенирование ДНК из костных фрагментов и зубов 1867 человек, живших в Евразии и Америке 31 630–150 лет назад, чтобы найти в них последовательности, сходные с геномом вируса натуральной оспы. Такие последовательности обнаружили у 13 индивидов. Как показали результаты радиоуглеродного датирования, 11 из них жили в 600–1050 гг. н.э. — период, включавший эпоху викингов. Для четырех из этих 11 реконструировали почти полные геномы вируса оспы. В результате генетическая история оспы стала длиннее на 1000 лет.
«Мы приняли участие в этом проекте, поскольку международная команда помимо генетического анализа населения эпохи викингов (основной проект, в котором работают ученые МГУ), решила проанализировать костные останки из тех регионов, где оспа была нередкой болезнью, и где викинги могли бывать, осваивая Северную Европу», –– сказала директор НИИ и музея антропологии МГУ академик Александра Бужилова, видный специалист в области палеопатологии.
В рамках исследования ученые МГУ работали с антропологической коллекцией Гнёздово – известного археологического комплекса X–XI веков, находящегося недалеко от современного Смоленска. Гнёздово было средневековым торговым центром, соединявшим Русь со странами Скандинавии (для одного из гнездовских образцов и была получена полная последовательность вирусного генома). Для обнаружения инфекции у народов Арктики, в частности, использовали коллекцию обдорских хантов XVIII-XIX вв. — оседлое население рыболовов, охотников и оленеводов из хранения Музея антропологии МГУ.
Одиннадцать древних последовательностей, восстановленных полностью или частично, попали в ныне вымершую кладу, сестринскую по отношению к более современным вирусам оспы, существовавшим до искоренения этой болезни, тогда как две последовательности XIX века группируются с современными. Предполагаемая дата существования последнего общего предка вирусов оспы человека с учетом новых данных —1700 лет назад.
У ортопоксвирусов (рода, к которому относится вирус оспы) количество функциональных генов обычно уменьшается при сужении диапазона хозяев. Действительно, геномы оспы эпохи викингов отличаются по этому признаку от современных. Три гена, активные во всех современных последовательностях, оказались неактивными во всех или некоторых древних геномах. В десяти генах, неактивных и в древних, и в современных геномах, инактивирующие мутации различны. Вероятно, у общего предка эти гены были активными, что предполагает параллельную эволюцию. Наконец, 14 генов, инактивированных в современных вирусах, активны в некоторых или во всех древних последовательностях, причем восемь из них кодируют известные факторы вирулентности или иммуномодуляторы. Таким образом, в процессе эволюции вируса оспы число активных генов явно снизилось.
Генетические данные, полученные в этом исследовании, в сочетании с ранними письменными данными об эпидемиях в южной и западной Европе говорят о том, что как минимум с конца VI века оспа на территории Европы присутствует повсеместно. Древние вирусы принадлежат к ранее неизвестной, ныне вымершей вирусной кладе, их эволюционный путь был иным, чем у современных вирусов. Различия в наборе активных генов, по-видимому, способствовали появлению вирусов оспы, способных широко распространиться в человеческой популяции, заключают авторы.
«Когда мы узнаем, как болезнь мутировала во времени, это дает нам возможность составить каталог мутаций, которые могут появиться в будущем: какие мутации и их комбинации делают такой патоген жизнеспособным и успешным? Если у них были эти мутации в прошлом, они, скорее всего, могут получить их снова. (…) Это один из немногих примеров, когда древние генетические исследования имеют прямое значение для современного и будущего здравоохранения», — говорит Мартин Сикора. Оспа побеждена, но в природе существуют родственные вирусы.
Источники
Barbara Mühlemann, et al. // Diverse variola virus (smallpox) strains were widespread in northern Europe in the Viking Age. // Science 24 Jul 2020: Vol. 369, Issue 6502, eaaw8977, DOI:
10.1126/science.aaw8977
Цитаты из пресс-релизов
Копенгагенского университета и МГУ
Источник