Вирус оспы тип его генома
Содержание статьи
еном вирусов оспы. Антигены вирусов оспы.
Геном вирусов оспы. Антигены вирусов оспы.Геном вирусов оспы представлен одной линейной молекулой двуцепочечной ДНК с ковалентно замкнутыми концами размером 130 тпн (парапоксвирусы) — 280 тпн (авипоксвирусы). На обоих концах генома имеются идентичные, но противоположно ориентированные тандемы повторяющихся нуклеотидных последовательностей. Геномы вирусов оспы способны кодировать около 200 белков, из которых не менее 100 входят в структуру вириона. Однако функциональные особенности определены лишь у небольшого количества вирусных белков. Наиболее важными из них являются ферменты, участвующие в синтезе вирусных нуклеиновых кислот и структурных компонентов вирионов. Например, синтез ДНК-полимеразы, ДНК-лигазы, РНК-полимеразы, энзимов, связанных с кэппированием и полиаденилированием мРНК и тимидинкиназы. Инфекционные вирусные частицы содержат системы транскрипции, которые могут выполнять in vitro синтез РНК, а также способны полиаденилировать, кэппировать и метилировать. В вирусных частицах содержится большое количество кодируемых вирусом энзимов и других биологически активных факторов. Некоторые гены вирусов оспы кодируют белки, которые секретируются инфицированными клетками и вызывают ответ организма на инфекцию, в том числе и формирование иммунитета. К таким вирокинам относятся гомологичный эпидермальный фактор роста, белок, снижающий активность комплемента, вирокины, обеспечивающие устойчивость к интерферону, и другие супрессоры иммунного ответа, подавляющие действие некоторых цитокинов организма хозяина. Вирусы оспы обычно характеризуются узким спектром хозяев. Они передаются чаще респираторным путем и реже через поврежденную кожу. Вирусы оспы овец, свиней, птиц и миксоматоза передаются также через укус членистоногими. Вирусы оспы устойчивы в окружающей среде и могут сохраняться годами в высохших струпьях кожи или других вируссодержащих материалах.
Большинство вирусов оспы хорошо размножаются в культуре клеток. Исключение составляют парапоксвирусы, вирус оспы свиней и вирус контагиозного моллюска. Однако они, так же как и ортопоксвирусы, легко образуют оспины на хориоал-лантоисной оболочке куриных эмбрионов. Вирусы оспы размножаются в цитоплазме, и, в отличие от других ДНК-вирусов, их размножение происходит независимо от ядра клетки, благодаря кодированию всех ферментов, необходимых для транскрипции и репликации вирусного генома. Некоторые из этих функций выполняются вирионами как таковыми. После слияния оболочки вириона с плазматической мембраной клетки или после эндоцитоза вирусная сердцевина освобождается в цитоплазму. Транскрипция вирусного генома характеризуется каскадностью, когда каждый временной класс генов («ранние», «промежуточные» и «поздние» гены) требует наличия специфических транскрипционных факторов, которые создаются предшествующим временным классом генов. Факторы, обеспечивающие транскрипцию промежуточных генов, кодируются ранними генами, тогда как факторы транскрипции поздних генов кодируются промежуточными генами. Транскрипция начинается вирионной транскриптазой и другими факторами, находящимися в сердцевине вириона, которые способны образовывать мРНК спустя минуты после инфицирования. Белки, образующиеся в результате трансляции этих мРНК, включая ДНК-полимеразу, тимидинкиназу и несколько других ферментов, необходимы для репликации вирусной ДНК. Репликация ДНК ВО связана с синтезом конкатемерных промежуточных структур, которые затем разрезаются с образованием единиц геномной длины. Детали этого процесса недостаточно изучены. С началом репликации ДНК происходит резкий сдвиг в генной экспрессии. Транскрипция «промежуточных» и «поздних» генов контролируется специфическими вирусными белками. Некоторые продукты транскрипции ранних генов образуются на поздней стадии инфекции, упаковываются в вири-оны и используются в следующем круге инфекции. Так как в состав вирусов оспы входит большое количество белков, не является неожиданным, что сборка вирионов есть комплексный процесс, который длится несколько часов и все еще целиком не выяснен. Образование вириона связано с вхождением ДНК внутрь незрелой сердцевинной структуры, которое затем завершается включением наружных покрывающих слоев. Репликация и сборка вирионов происходят в разных местах цитоплазмы в так называемых виропластах или вирусных фабриках. Вирионы выходят из клетки почкованием (оболочечные вирионы), или путем экзоцитоза, или при лизисе клеток (вирионы без оболочки). Большинство вирионов освобождаются при цитолизе и не имеют оболочки. Вирионы с оболочкой и без нее обладают инфекционностью, но первые, вероятно, играют более значительную роль в возникновении и распространении заболевания, а также в создании иммунитета. В очищенном вирусе осповакцины (ВОВ) выявлены белки с молекулярной массой 10-250 кД. Многие из них сосредоточены в сердцевине вириона. Два структурных гликопротеина располагаются между оболочкой и сердцевиной. В оболочке ВОВ содержится около 10 белков, из которых иммунологически наиболее активны крупномолекулярные белки с молекулярной массой 58-32 кД (VP4c, VP6a, VP6b и VP7a). Белок 32 кД определяет круг хозяев и важен для репликации вируса. В составе очищенного вируса оспы птиц обнаружено 29 полипептидов с молекулярной массой 14-138 кД. Наивысшей антигенной и иммуногенной активностью обладают полипептиды с молекулярной массой 35 и 37 кД. За индукцию вируснейтрализующих антител ВО ответственны антигены, расположенные на поверхности наружной оболочки вириона, и прежде всего белок 58 кД (VP4c), являющийся основным структурным компонентом трубочек (ворсинок). Антисыворотка к этому белку нейтрализовала инфекционность вируса и предотвращала образование синцития в культуре клеток. Этот белок ответственен за выработку иммунитета. Внеклеточные вирионы покрыты дополнительной наружной оболочкой, отсутствующей у внутриклеточных вирионов. Она играет важную роль в индукции синтеза ВН-антител. Инфекционность ВОВ и ВО крупного рогатого скота, имеющих наружную оболочку, нейтрализовалась антисывороткой к имеющему эту оболочку ВОВ, но не нейтрализовалась антисывороткой к ВОВ, лишенному наружной оболочки. За гемагглютинирующую активность ортопоксвирусов ответственны полипептид 85 кД и гликопротеид 41 кД. Во внеклеточном оболочечном ВОВ гемагглютинирующие свойства связаны с полипептидом 85 кД. Внутриклеточные вирионы (без дополнительной оболочки) практически не содержали этого полипептида. Неструктурный гемагглютинин формируется на цитоплазматических мембранах. С его образованием инфицированные клетки приобретают способность адсорбировать эритроциты. Вирусспецифические белки с молекулярной массой 32 и 37 кД, экспрессируемые на поверхности клеток, инфицированных ВОВ, делают их мишенями для специфических цитотоксических Т-лимфоцитов. МАТ-реактивные против каждого из пяти (54; 34; 32; 29 и 17-25 кД) белков наружного слоя поверхности вируса осповакцины нейтрализовали его инфекционность. В структуре полипептида 54 кД обнаружено два нейтрализующих эпитопа(Аи В). Анализ антигенных детерминант поверхностных полипептидов, проведенный с помощью МАТ, выявил в составе ортопоксвирусов, наряду с видоспецифическими, группоспецифические эпитопы. Связывание ВОВ с нейтрализующими МАТ не препятствовало его прикреплению к клеткам-мишеням, но блокировало депротеинизацию вирионов. — Также рекомендуем «Оспа овец. Оспа птиц. Особенности оспы овец и птиц.» Оглавление темы «Частная вирусология.»: 1. ДНК-вакцины. Свойства и методы получения днк-вакцин. 2. Синтетические пептидные вакцины. Свойства синтетических пептидных вакцин. 3. Вирусы оспы. Виды вирусов оспы. Строение вирусов оспы. 4. Геном вирусов оспы. Антигены вирусов оспы. 5. Оспа овец. Оспа птиц. Особенности оспы овец и птиц. 6. Контагиозный пустулёзный дерматит. Эктима. Миксоматоз. 7. Асфаровирусы. Признаки и свойства асфаровирусов. 8. Иридовирусы. Герпесвирусы. Семейство герпесвирусов. 9. Эпидемиология герпесвирусов. Строение герпесвирусов. 10. Антигены вируса герпеса. Протеины герпесвирусов. |
Источник
Poxviridae — Poxviridae
Семейство вирусов
Poxviridae — это семейство вирусов . Люди, позвоночные и членистоногие служат естественными хозяевами. В настоящее время в этом семействе 83 вида, разделенных на 22 рода, которые делятся на два подсемейства. Болезни, связанные с этим семейством, включают оспу .
Четыре рода поксвирусов могут инфицировать людей: ортопоксвирус , парапоксвирус , ятапоксвирус , моллюскципоксвирус . Orthopox : вирус оспы (натуральная оспа), вирус осповакцины , вирус коровьей оспы , вирус оспы обезьян ; Parapox : orf вирус, pseudocowpox , вирус папулезного стоматита крупного рогатого скота ; Yatapox : вирус танапокса , вирус опухоли обезьяны yaba ; Molluscipox : вирус контагиозного моллюска (MCV). Наиболее распространены осповакцина (встречается на Индийском субконтиненте) и контагиозный моллюск, но число случаев заражения оспой обезьян растет (наблюдается в странах с тропическими лесами Западной и Центральной Африки). Заболевание с аналогичным названием ветряная оспа не является истинным поксвирусом и на самом деле вызвано вирусом герпеса ветряной оспой .
Филогения
A новая систематика была предложена недавно после открытия нового поксвируса белки в Берлине, Германия.
Структура
A) Электронная микрофотография частиц поксвируса в синовиальной оболочке большой коричневой летучей мыши на северо-западе США. Б) Отрицательное окрашивание частиц поксвируса в супернатанте клеточной культуры. Масштабная линейка = 100 нм.
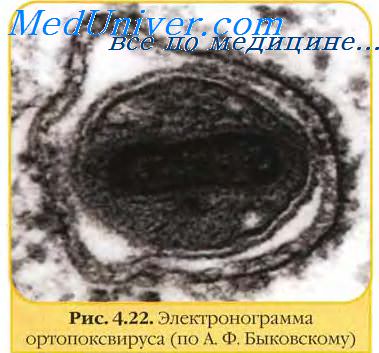
Вирусные частицы (вирионы) Poxviridae обычно покрыты оболочкой (вирион с внешней оболочкой), хотя внутриклеточная зрелая форма вириона вируса, которая содержит другую оболочку , также является заразным. Они различаются по своей форме в зависимости от вида, но обычно имеют форму кирпича или овальную форму, похожую на округлый кирпич, потому что они окружены эндоплазматической сетью. Вирион исключительно большой, его размер составляет около 200 нм в диаметре и 300 нм в длину и несет его геном в виде одноцепочечной, линейной, двухцепочечной сегмент ДНК. Для сравнения: риновирус в 10 раз меньше типичного вириона Poxviridae.
Репликация
Репликация поксвируса включает несколько стадий. Сначала вирус связывается с рецептором на поверхности клетки-хозяина; Считается, что рецепторами поксвируса являются гликозаминогликаны . После связывания с рецептором вирус проникает в клетку, где он обнажается. Удаление оболочки вируса — это двухэтапный процесс. Сначала удаляется внешняя мембрана, когда частица попадает в клетку; во-вторых, вирусная частица (без внешней мембраны) сливается с клеточной мембраной, высвобождая ядро в цитоплазму. Гены вируса оспы экспрессируются в две фазы. Ранние гены кодируют неструктурный белок, включая белки, необходимые для репликации вирусного генома, и экспрессируются перед репликацией генома. Поздние гены экспрессируются после репликации генома и кодируют структурные белки для создания вирусной частицы. Сборка вирусной частицы происходит за пять стадий созревания, которые приводят к окончательному экзоцитозу нового вириона, заключенного в оболочку. После репликации генома незрелый вирион собирает белок A5 для создания внутриклеточного зрелого вириона. Белок выравнивается и кирпичная оболочка внутриклеточного обволакивающего вириона. Затем эти частицы сливаются с клеточной плазмой с образованием связанного с клеткой оболочечного вириона, который встречается с микротрубочками и готовится покинуть клетку в виде внеклеточного оболочечного вириона. Сборка вирусной частицы происходит в цитоплазме клетки и представляет собой сложный процесс, который в настоящее время исследуется для более глубокого понимания каждой стадии. Учитывая тот факт, что этот вирус большой и сложный, репликация происходит относительно быстро и занимает около 12 часов, пока клетка-хозяин не погибнет в результате высвобождения вирусов.
Репликация поксвируса необычна для вируса с двухцепочечным геномом ДНК , поскольку она происходит в цитоплазме, хотя это типично для других крупных ДНК-вирусов. Поксвирус кодирует собственный механизм транскрипции генома, ДНК-зависимую РНК-полимеразу, которая делает возможной репликацию в цитоплазме. Большинству двухцепочечных ДНК-вирусов требуется ДНК-зависимая РНК-полимераза клетки-хозяина для выполнения транскрипции. Эти ДНК-хозяева находятся в ядре , и поэтому большинство вирусов с двухцепочечной ДНК проводят часть своего инфекционного цикла в ядре клетки-хозяина.
Эволюция
Предок поксвирусов неизвестен, но структурные исследования предполагают, что это мог быть аденовирус или вид, связанный как с поксвирусами, так и с аденовирусами.
На основании Что касается организации генома и механизма репликации ДНК, кажется, что филогенетические отношения могут существовать между рудивирусами (Rudiviridae ) и крупными эукариальными ДНК-вирусами: вирусом африканской чумы свиней (Asfarviridae ), хлореллой вирусов (Phycodiridae ) и поксвирусов (Poxviridae).
Частота мутаций в этих геномах оценивается в 0,9-1,2 x 10 замен на сайт в год. Согласно второй оценке, этот показатель составляет 0,5-7 × 10 замен нуклеотидов на сайт в год. Третья оценка показывает, что этот показатель составляет 4-6 × 10.
Последний общий предок существующих поксвирусов, поражающих позвоночных, существовал 0,5 миллионов лет назад. Род Avipoxvirus отошел от своего предка 249 ± 69 тысяч лет назад. Предок рода Orthopoxvirus должен был отличиться от других клад 0,3 миллиона лет назад. Согласно второй оценке времени расхождения, это событие произошло 166 000 ± 43 000 лет назад. Разделение Orthopox на дошедшие до нас роды произошло около 14000 лет назад. Род Leporipoxvirus разошелся ~ 137 000 ± 35 000 лет назад. Затем последовал предок рода Yatapoxvirus. Последний общий предок Capripoxvirus и Suipoxvirus разошелся 111 000 ± 29 000 лет назад.
Изолят рыб — поксвирус лососевых жабр — по всей видимости, является самой ранней ветвью Chordopoxvirinae.
Оспа
Дата появления оспы не установлена. Скорее всего, он произошел от вируса грызунов между 68 000 и 16 000 лет назад. Широкий диапазон дат связан с разными записями, используемыми для калибровки молекулярных часов. Одной из кладов был штамм натуральной оспы (более тяжелая форма оспы), распространившийся из Азии между 400 и 1600 годами назад. Вторая клада включала как аластрим минор (фенотипически легкую оспу), описанный с американских континентов, так и изоляты из Западной Африки, которые произошли от предкового штамма между 1400 и 6300 годами ранее. Эта клада далее разделилась на две субклада, по крайней мере, 800 лет назад.
Согласно второй оценке, натуральная оспа была выделена 3000-4000 лет назад. Это согласуется с археологическими и историческими данными о появлении оспы как болезни человека, которые предполагают относительно недавнее происхождение. Однако, если предположить, что частота мутаций аналогична частоте мутаций герпесвирусов , дата расхождения между натуральной оспой и татерапоксом оценивается как 50 000 лет назад. Хотя это согласуется с другими опубликованными оценками, это предполагает, что археологические и исторические свидетельства очень неполны. Необходимы более точные оценки частоты мутаций в этих вирусах.
Таксономия
Выделяются следующие подсемейства и роды (-virinae обозначает подсемейство, а -virus обозначает род):
- Chordopoxvirinae
- Avipoxvirus
- Capripoxvirus
- Cervidpoxvirus
- Crocodylidpoxvirus
- Leporipoxvirus
- Molluscipoxvirus
- Orthopoxvirus
- Parapoxvirus
- Suipoxvirus
- Yatapoxvirus
- Entomopoxvirinae
- Alphaentomopoxvirus
- Entomopoxvirinae
- Alphaentomopoxvirus
- The Familypoxvirus
- The Poxviridae — это наследие исходной группы вирусов, связанных с заболеваниями, вызывающими оспу на коже. Современная классификация вирусов основана на фенотипических характеристиках; морфология, тип нуклеиновой кислоты, способ репликации, организмы-хозяева и тип заболевания, которое они вызывают. Вирус оспы остается наиболее заметным членом этого семейства.
Виды подсемейства Chordopoxvirinae инфицируют позвоночных , а виды подсемейства Entomopoxvirinae заражают насекомых . Существует 10 признанных родов Chordopoxvirinae и 3 — Entomopoxvirinae. Оба подсемейства также содержат ряд неклассифицированных видов, для которых в будущем могут быть созданы новые роды. Вирус котия — необычный вирус, который может принадлежать к новому роду. Еще два поксвируса — NY_014 и Мурманский поксвирус.
Примечания
GC-содержание этих геномов значительно различается. Avipoxvirus, Capripoxvirus, Cervidpoxvirus, Orthopoxvirus, Suipoxvirus, Yatapoxvirus и один род Entomopox (Betaentomopoxvirus), а также несколько других неклассифицированных Entomopoxvirus имеют низкое содержание G + C, в то время как другие — Molluscipoxvirus, Paraopoxvirus и Paraopoxvirus относительно высокое, Paraopoxvirus и Paraopoxvirus. C содержание. Причины этих различий неизвестны.
Филогенетический анализ геномов 26 хордопоксвирусов показал, что центральная область генома консервативна и содержит ~ 90 генов. Концы, напротив, не сохраняются у разных видов. Из этой группы наиболее расходится Avipoxvirus. Следующим по размеру является Molluscipoxvirus. Роды Capripoxvirus, Leporipoxvirus, Suipoxvirus и Yatapoxvirus объединяются в кластер: Capripoxvirus и Suipoxvirus имеют общего предка и отличаются от рода Orthopoxvirus. В пределах рода Othopoxvirus штамм вируса коровьей оспы Brighton Red, вирус эктромелии и вирус оспы обезьян не группируются близко ни к одному другому представителю. Вирус натуральной оспы и вирус оспы верблюда образуют подгруппу. Вирус осповакцины наиболее близок к CPV-GRI-90.
Вирус коровьей оспы
Прототипом поксвируса является вирус осповакцины , известный своей ролью в искоренении оспы. Вирус коровьей оспы является эффективным инструментом для экспрессии чужеродного белка, поскольку он вызывает сильный иммунный ответ хозяина. Вирус осповакцины проникает в клетки в основном путем слияния клеток, хотя в настоящее время ответственный за него рецептор неизвестен.
Вакцина содержит три класса генов: ранние, промежуточные и поздние. Эти гены транскрибируются вирусной РНК-полимеразой и соответствующими факторами транскрипции. Вакцина реплицирует свой геном в цитоплазме инфицированных клеток, и после поздней стадии экспрессии гена претерпевает морфогенез вириона, который производит внутриклеточные зрелые вирионы, содержащиеся внутри оболочки оболочки. Происхождение оболочки оболочки до сих пор неизвестно. Затем внутриклеточные зрелые вирионы транспортируются в аппарат Гольджи, где они обертываются двумя дополнительными мембранами, становясь вирусом с внутриклеточной оболочкой. Он транспортируется по микротрубочкам цитоскелета, чтобы достичь периферии клетки, где он сливается с плазматической мембраной, становясь связанным с клеткой оболочечным вирусом. Это запускает актиновые хвосты на поверхности клеток или высвобождается в виде вириона, окруженного внешней оболочкой.
История
Заболевания, вызываемые вирусами оспы, особенно оспа, были известны на протяжении веков. Один из самых ранних подозреваемых случаев связан с египетским фараономРамзесом V , который, как полагают, умер от оспы около 1150 года до нашей эры. Считалось, что оспа была перенесена в Европу в начале 8-го века, а затем в Америку в начале 16-го века, в результате чего за два года после появления этого заболевания погибло 3,2 миллиона ацтеков. Такое количество погибших можно объяснить полным отсутствием воздействия вируса на американское население на протяжении тысячелетий. Спустя столетие после того, как Эдвард Дженнер показал, что менее сильнодействующая коровья оспа может быть использована для эффективной вакцинации против более смертоносной оспы, мировые усилия по вакцинации всех против оспы начались с конечной целью избавить мир от эпидемии, похожей на чуму. Последний случай эндемической оспы произошел в Сомали в 1977 году. В результате обширных поисков, продолжавшихся в течение двух лет, новых случаев не было обнаружено, и в 1979 году Всемирная организация здравоохранения (ВОЗ) объявила, что болезнь официально искоренена. В 1986 году все образцы вируса были уничтожены или переданы в две утвержденные справочные лаборатории ВОЗ: в штаб-квартире федеральных Центров по контролю и профилактике заболеваний (CDC) в Атланте , Джорджия ( США) и в Институте вирусных препаратов в Москве. После 11 сентября 2001 года правительства США и Великобритании стали проявлять повышенную озабоченность по поводу использования натуральной оспы или заболевания, похожего на оспу, в биотерроризме.
Ссылки
Внешние ссылки
- Электронные микрофотографии ортопоксвирусов и парапоксвирусов родов , включая вирус натуральной оспы , были собраны Международный комитет по таксономии вирусов в своей картинной галерее Poxviridae .
- Buller, R. Mark L .; Паламбо, Грегори Дж. (1991). «Патогенез поксвируса» . Обзоры микробиологии и молекулярной биологии. 55 (1): 80-122. PMC372802 . PMID1851533 .
- Страница таксономии NCBI .
- Poxviridae в Центре ресурсов вирусной биоинформатики .
- Viralzone : Poxviridae
- ICTV
- База данных и анализ вирусных патогенов ( ViPR): Poxviridae
Источник