Вирус натуральной оспы симметрия
Содержание статьи
ирусы. Вирион. Морфология вирусов. Размеры вирусов. Нуклеиновые кислоты вирусов.
Оглавление темы «Типы микроорганизмов. Вирусы. Вирион.»: Вирусы. Вирион. Морфология вирусов. Размеры вирусов. Нуклеиновые кислоты вирусов.Вирусы [от лат. virus, яд] — наименьшие по размерам агенты, имеющие геном, окружённый белковой оболочкой. Вирусы не воспроизводятся самостоятельно, они — облигатные внутриклеточные паразиты, репродуцирующиеся только в живых клетках. Все вирусы существуют в двух формах. В настоящее время известны вирусы бактерий (бактериофаги), грибов, растений и животных. Внеклеточная форма — вирион — включает в себя все составные элементы (капсид, нуклеиновую кислоту, структурные белки, ферменты и др.). Внутриклеточная форма — вирус — может быть представлена лишь одной молекулой нуклеиновой кислоты, так как, попадая в клетку, вирион распадается на составные элементы.
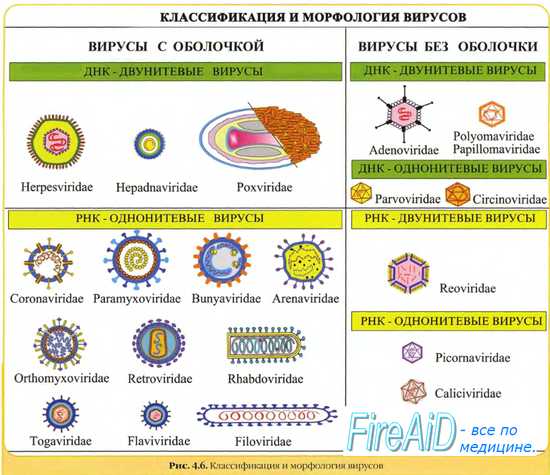
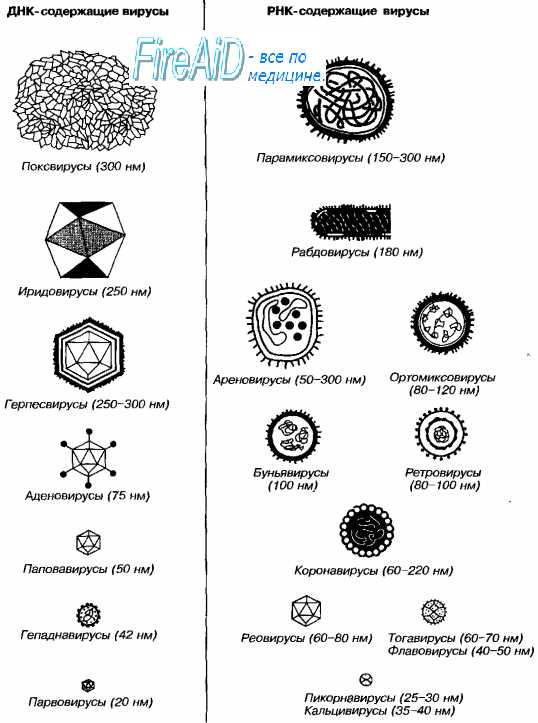
Морфология вирусов. Размеры вирусов.Несмотря на внутриклеточный паразитизм, среди вирусов имеются крупные виды, соизмеримые по размерам с микоплазмами и хламидиями. Например, вирус натуральной оспы достигает 400 нм и вполне сравним с риккетсиями (300-500 нм) и хламидиями (300-400 нм). По морфологии выделяют вирусы палочковидные (например, возбудитель лихорадки Эбола), пуле-видные (вирус бешенства), сферические (герпесвирусы), овальные (вирус оспы), а также бактериофаги, имеющие сложную форму (рис. 2-1). При всём разнообразии конфигураций, размеров и функциональных характеристик вирусам присущи некоторые общие признаки. В общем виде зрелая вирусная частица (вирион) состоит из нуклеиновой кислоты, белков и липидов, либо в его состав входят только нуклеиновые кислоты и белки. Нуклеиновые кислоты вирусовВирусы содержат только один тип нуклеиновой кислоты, ДИК или РНК, но не оба типа одновременно. Например, вирусы оспы, простого герпеса, Эпстайна-Барр — ДНК-содержащие, а тогавирусы, пикорнавирусы — РНК-содержащие. Геном вирусной частицы гаплоидный. Наиболее простой вирусный геном кодирует 3-4 белка, наиболее сложный — более 50 полипептидов. Нуклеиновые кислоты представлены однонитевыми молекулами РНК (исключая реовиру-сы, у которых геном образован двумя нитями РНК) или двухнитевыми молекулами ДНК (исключая парвовирусы, у которых геном образован одной нитью ДНК). У вируса гепатита В нити двухнитевой молекулы ДНК неодинаковы по длине. Вирусные ДНК образуют циркулярные, ковалентно-сцёпленные суперспирализованные (например, у паповавирусов) или линейные двухнитевые структуры (например, у герпес- и аденовирусов). Их молекулярная масса в 10-100 раз меньше массы бактериальных ДНК. Транскрипция вирусной ДНК (синтез мРНК) осуществляется в ядре заражённой вирусом клетки. В вирусной ДНК на концах молекулы имеются прямые или инвертированные (развёрнутые на 180″) повторяющиеся нуклеотидные последовательности. Их наличие обеспечивает способность молекулы ДНК замыкаться в кольцо. Эти последовательности, присутствующие в одно- и двух-нитевых молекулах ДНК, — своеобразные маркёры вирусной ДНК.
Вирусные РНК представлены одно- или двухнитевыми молекулами. Однонитевые молекулы могут быть сегментированными — от 2 сегментов у ареновирусов до 11 — у ротавирусов. Наличие сегментов ведёт к увеличению кодирующей ёмкости генома. Вирусные РНК подразделяют на следующие группы: плюс-нити РНК (+РНК), минус-нити РНК (-РНК). У различных вирусов геном могут образовывать нити +РНК либо -РНК, а также двойные нити, одна из которых -РНК, другая (комплементарная ей) — +РНК. Плюс-нити РНК представлены одиночными цепочками, имеющими характерные окончания («шапочки») для распознавания рибосом. К этой группе относят РНК, способные непосредственно транслировать генетическую информацию на рибосомах заражённой вирусом клетки, то есть выполнять функции мРНК. Плюс-нити выполняют следующие функции: служат мРНК для синтеза структурных белков, матрицей для репликации РНК, упаковываются в капсид с образованием дочерней популяции. Минус-нити РНК не способны транслировать генетическую информацию непосредственно на рибосомах, то есть они не могут функционировать как мРНК. Однако такие РНК служат матрицей для синтеза мРНК. Инфекционность нуклеиновых кислот вирусовМногие вирусные нуклеиновые кислоты инфекционны сами по себе, так как содержат всю генетическую информацию, необходимую для синтеза новых вирусных частиц. Эта информация реализуется после проникновения вириона в чувствительную клетку. Инфекционные свойства проявляют нуклеиновые кислоты большинства +РНК- и ДНК-содержащих вирусов. Двухнитевые РНК и большинство -РНК не проявляют инфекционных свойств. — Также рекомендуем «Капсид вируса. Функции капсида вирусов. Капсомеры. Нуклеокапсид вирусов. Спиральная симметрия нуклеокапсида. Кубическая симметрия капсида.» |
Источник
Тип симметрии вируса натуральной оспы
Типы симметрии вирусов
Капсомеры вирионаорганизованы в один или два слоя по двум типам симметрии – кубическому или спиральному (рисунок 4). Симметричность капсида связана с тем, что для упаковки генома требуется большое количество капсомеров, а компактное их соединение возможно лишь при условии симметричного расположения субъединиц. Формирование капсида напоминает процесс кристаллизации и протекает по принципу самосборки.
Рисунок 4 – Основные типы симметрии вирусов:
А– кубический («голый» вирион); Б – кубический («одетый» вирион); В – спиральный («голый» вирион); Г – спиральный («одетый» вирион)
Спиральная симметрия.В нуклеокапсиде взаимодействие нуклеиновой кислоты и белка осуществляется по одной оси вращения. Каждый вирус со спиральной симметрией обладает характерной длиной, шириной и периодичностью нуклеокапсида. Нуклеокапсиды большинства патогенных для человека вирусов имеют спиральную симметрию. К этой группе относят и вирус табачной мозаики (рисунок 5), капсид которого образован 2130 одинаковыми белковыми субъединицами. Организация по принципу спиральной симметрии придаёт вирусам палочковидную форму. При спиральной симметрии белковый чехол лучше защищает наследственную информацию, но требует большого количества белка, так как покрытие состоит из сравнительно крупных блоков.
Рисунок 6 – Один из возможных вариантов кубической симметрии |
Рисунок 5 – Вирус табачной мозаики (спиральный тип симметрии) |
Кубическая симметрия.У подобных вирусов нуклеиновая кислота окружена капсомерами, образующими фигуру икосаэдра – многогранника с 12 вершинами, 20 треугольными гранями и 30 углами (рисунок 6). Организация по принципу кубической симметрии придаёт вирусам сферическую форму. Принцип кубической симметрии – самый экономичный для формирования замкнутого капсида, так как для его организации используются сравнительно небольшие белковые блоки, образующие большое внутреннее пространство, в которое свободно укладывается нуклеиновая кислота.
Двойная (смешанная) симметрия.Некоторые бактериофаги (вирусы бактерий) имеют двойную симметрию: головка организована по принципу кубической симметрии, отросток – по принципу спиральной симметрии (рисунок 7).
Отсутствие постоянной симметрии.Для вирусов больших размеров (для поксвирусов) характерно отсутствие постоянной симметрии.
Не нашли то, что искали? Воспользуйтесь поиском:
Лучшие изречения: Как то на паре, один преподаватель сказал, когда лекция заканчивалась – это был конец пары: “Что-то тут концом пахнет”. 8752 –
| 8286 – или читать все.
источник
1)Строение (структура, химический состав) вирусов. Типы симметрии нуклеокапсида.
Вирусы— мельчайшие микробы, не имеющие клеточного строения, белоксинтезирующей системы, содержащие только ДНК или РНК. Относятся к царству Vira. Являясь облигатными внутриклеточными паразитами, вирусы размножаются в цитоплазме или ядре клетки. Они — автономные генетические структуры. Отличаются особым — разобщенным (дизъюнктивным) способом размножения (репродукции): в клетке отдельно синтезируются нуклеиновые кислоты вирусов и их белки, затем происходит их сборка в вирусные частицы. Сформированная вирусная частица называется вирионом.
Морфологию и структурувирусов изучают с помощью электронного микроскопа, так как их размеры малы и сравнимы с толщиной оболочки бактерий.
Формавирионов может быть различной: палочковидной (вирус табачной мозаики), пулевидной (вирус бешенства), сферической (вирусы полиомиелита, ВИЧ), в виде сперматозоида (многие бактериофаги). Различают просто устроенные и сложно устроенные вирусы.
Простые, или безоболочечные, вирусы состоят из нуклеиновой кислоты и белковой оболочки, называемой капсидом. Капсид состоит из повторяющихся морфологических субъединиц — капсомеров. Нуклеиновая кислота и капсид взаимодействуют друг с другом, образуя нуклеокапсид.
Сложные, или оболочечные, вирусы снаружи капсида окружены липопротеиновой оболочкой (суперкапсидом, или пеплосом). Эта оболочка является производной структурой от мембран вирус-инфицированной клетки. На оболочке вируса расположены гликопротеиновые шипы, или шипики (пепломеры). Под оболочкой некоторых вирусов находится матриксный М-белок.
Фенотипическое смешивание довольно широко распространено среди близкородственных безоболочечных вирусов, таких, например, как вирусы полиомиелита типов 1 и 2, вирусов ЭКХО и Коксаки, других пикорнавирусов. Таким образом, немутационные вирусы-гибриды – полноценные вирионы. Подобно вирусам-мутантам, возникают путем комплементации, а не вследствие скрещивания геномов, как рекомбинанты. Состояния гетерозиготности и транскапсидизации вирусов неустойчивы и быстро исчезают при пассажах. Биологическое значение гетерозигот не выяснено. Транскапсидизация же может обеспечить вирусам-гибридам широкий круг хозяев и преодоление межвидовых барьеров.
Капсид и суперкапсидзащищают вирионы от влияния окружающей среды, обусловливают избирательное взаимодействие (адсорбцию) с клетками, определяют антигенные и иммуногенные свойства вирионов. Внутренние структуры вирусов называются сердцевиной.
Тип симметрии. Капсид или нуклеокапсид могут иметь спиральный, икосаэдрический (кубический) или сложный тип симметрии.Икосаэдрическийтип симметрии обусловлен образованием изометрически полого тела из капсида, содержащего вирусную нуклеиновую кислоту (например, у вирусов гепатита А, герпеса, полиомиелита).Спиральныйтип симметрии обусловлен винтообразной структурой нуклеокапсида (например, у вируса гриппа).
Включения— скопление вирионов или отдельных их компонентов в цитоплазме или ядре клеток, выявляемые под микроскопом при специальном окрашивании. Вирус натуральной оспы образует цитоплазматические включения — тельца Гварниери; вирусы герпеса и аденовирусы — внутриядерные включения.
Размеры вирусов определяют с помощью электронной микроскопии, методом ультрафильтрации через фильтры с известным диаметром пор, методом ультрацентрифугирования. Одним из самых мелких вирусов является вирус полиомиелита (около 20 нм), наиболее крупным — натуральной оспы (около 350 нм).
Вирусы имеют уникальный геном, так как содержат либо ДНК, либо РНК. Поэтому различают ДНК-содержащие и РНК-содержащие вирусы. Они обычно гаплоидны, т.е. имеют один набор генов. Геном вирусов представлен различными видами нуклеиновых кислот: двунитчатыми, однонитчатыми, линейными, кольцевыми, фрагментированными. Среди РНК-содержащих вирусов различают вирусы с положительным (плюс-нить РНК) геномом. Плюс-нить РНК этих вирусов выполняет наследственную функцию и функцию информационной РНК (иРНК). Имеются также РНК-содержащие вирусы с отрицательным (минус-нить РНК) геномом. Минус-нить РНК этих вирусов выполняет только наследственную функцию.
Геномвирусов способен включаться в состав генетического аппарата клетки в виде провируса, проявляя себя генетическим паразитом клетки. Нуклеиновые кислоты некоторых вирусов (вирусы герпеса и др.) могут находиться в цитоплазме инфицированных клеток, напоминая плазмиды.
Внутримолекулярные рекомбинации у вирусов, как и других микроорганизмов, реализуются механизмом разрыв – воссоединение, а у РНК-вирусов с сегментированным геномом – перемешиванием генов. В общем, у вирусов различают две группы рекомбинаций -рекомбинация у ДНК-вирусов и рекомбинация-пересортировка у РНК-вирусов с сегментированным геномом.
Среди генетических рекомбинаций ДНК-вирусов выделяют рекомбинации:
1) между двумя дикими типами вирусов с интактными (лат intactus – нетронутый), т. е. полными, геномами;
2) между диким типом и его мутантным вариантом;
3) между вариантами мутантов дикого типа вируса.
источник
Методы лабораторной диагностики натуральной оспы
Сущность натуральной оспы.
Назовите формы натуральной оспы в зависимости от клинических симптомов.
Особенности иммунитета, который формируется у пациента, переболевшего натуральной оспой.
Механизмы и пути инфицирования человека возбудителем натуральной оспы.
Особенности морфологии возбудителя натуральной оспы.
Структура генома вируса, вызывающего натуральную оспу.
Форма вируса натуральной оспы.
Укажите систематическое положение возбудителя натуральной оспы.
Натуральная оспа
+ Вирион имеет кирпичеобразную форму с закругленными концами или овоидную форму.
+ Двунитевая нефрагментированная ДНК.
+ Сердцевина вируса имеет вид двояковогнутого диска.
+ Внутренняя гладкая мембрана окружает сердцевину и латеральные тельца.
+ Внешняя мембрана имеет трубчатые структуры.
*** в клетке-хозяине, репродукция вируса происходит в цитоплазме.
*** антигенная структура вируса, который является возбудителем натуральной оспы.
+ Нуклеопротеин (NP) – группоспецифический антиген.
*** входные ворота для возбудителя натуральной оспы.
+ Верхние дыхательные пути и кожа.
*** вирусемия при заболевании пациента натуральной оспой характерна.
+ Аэрогенный (воздушно-капельный, воздушно-пылевой).
+ Контактный путь: через вещи больного, содержимое оспенных везикул, частицы струпа.
+ Напряженный на протяжении всей жизни гуморальный.
+ В данное время иммунная защита у людей отсутствует.
+ Очень тяжелая: геморрагическая или “черная оспа” (характеризуется кровоизлияниями в кожу и кровотечениями).
+ Очень тяжелая: оспенная пурпура или “красная оспа” (еще до появления сыпи характерны мощные кровотечения, сильные кровоизлияния под кожу).
+ Тяжелая – сливающаяся оспа.
+ Легкая форма – алястрим [variola minor] (оспа без сыпи и лихорадки)
+ Вариолоид – своеобразное развитие оспы у вакцинированных лиц.
+ Особоопасная высококонтагиозная конвенционная экзогенная моноциклическая вирусная инфекция, характеризующаяся тяжелым течением, лихорадкой, интоксикацией, мощной пустуллезно-папулезной сыпью на коже и слизистых оболочках, формированием струпов рубцов после отпадания струпов.
Дата добавления: 2014-10-31 ; Просмотров: 497 ; Нарушение авторских прав?
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
источник
Тип симметрии вируса натуральной оспы
Натуральная оспа (variola vera) — особо опасная вирусная инфекция, проявляющаяся интоксикацией, лихорадкой и пустулёзно-папулёзной сыпью.
Оспа одна из древнейших инфекционных болезней человека. Первые упоминания об натуральной оспе относятся к 3730-3710 гг. до н.э. (папирус Аменофиса I). В Европу возбудитель проник в VI веке, в Россию был занесён в XV-XVI веках, в Америку — в XVI веке.
До открытия Дженнером метода вакцинации европейские врачи были бессильны перед натуральной оспой. В отдельные годы смертность от оспы достигала 1,5 млн человек.
Ещё в 1974 г. в Индии было отмечено 31 262 летальных исхода натуральной оспы. Последний случай заболевания, зарегистрирован в 1977 г. в Сомали, а через несколько лет ВОЗ объявила о ликвидации натуральной оспы.
Возбудитель оспы — самый крупный вирус (220×300 нм), имеет кирпичеобразную форму с закруглёнными углами.
В состав вируса натуральной оспы входят 30 белков, более 10 из них — ферменты, катализирующие синтез нуклеиновых кислот. Вирус натуральной оспы проявляет гемагглютинирующие свойства; гемагглютинин состоит из трёх гликопротеинов.
Важнейшие Аг вируса оспы — нуклеопротеид NP (общий для всего семейства), термостабильный и термолабильный Аг, а также группа растворимых Аг. Размеры вирионов позволяют обнаружить их световой микроскопией в форме телец включений.
Внутриклеточные (точнее околоядерные) включения в клетках роговицы заражённого кролика впервые обнаружил итальянский патолог Д. Гварнери (1892). ЭТ возбудителя натуральной оспы в отделяемом везикул открыл немецкий бактериолог Э. Пашен (1906). Позднее было установлено наличие двух штаммов вируса натуральной оспы.
Первый вызывает классическую оспу (variola major) с летальностью, превышающей 50%, второй — алястрим (variola minor) с более лёгким течением и летальностью, не превышающей 1%. Возбудители идентичны по основным свойствам. Отличительные особенности — способность вируса алястрима размножаться в куриных эмбрионах при более низкой температуре (37,5 °С), образовывать бляшки и проявлять цитопатичес-кий эффект в культуре куриных фибробластов при температуре 37 °С.
К вирусу натуральной оспы чувствителен человек и приматы; экспериментальное внутримозговое заражение новорождённых мышат приводит к развитию генерализованной инфекции. Взрослые мыши нечувствительны к вирусу.
источник
Вирус натуральной оспы
Вирус вызывает особо опасное высококонтагиозное инфекционное заболевание, характеризующееся общим поражением организма и обильной сыпью на коже и слизистых оболочках. В прошлом отмечались эпидемии и пандемии заболевания, сопровождающиеся высокой летальностью. В 1892 г. Г.Гварниери, исследуя под микроскопом срезы роговицы зараженного кролика, обнаружил специфические включения, впоследствии названные тельцами Гварниери, представляющие собой скопления вирусов натуральной оспы. Возбудитель оспы впервые обнаружен в световом микроскопе Е. Пашеном (1906).
Таксономия. Вирус натуральной оспы – ДНК-содержащий; относится к семейству Poxviridae (от англ, рох – язва), роду Orthopoxvirus.
Морфология, химический состав, антигенная структура. Вирус натуральной оспы является самым крупным вирусом, при электронной микроскопии имеет кирпичеобразную форму с закругленными углами размером 250-400 нм. Вирион состоит из сердцевины, имеющей форму гантели, двух боковых тел, расположенных по обе стороны от сердцевины, трехслойной наружной оболочки. Вирус содержит линейную двунитчатую ДНК, более 30 структурных белков, включая ферменты, а также липиды и углеводы.В составе вируса обнаружено несколько антигенов: нуклео-протеидный, растворимые и гемагглютинин. Вирус натуральной оспы имеет общие антигены с вирусом осповакцины (коровьейоспы).
Культивирование. Вирусы хорошо размножаются в куриных эмбрионах, образуя белые плотные бляшки на хорионаллантоисной оболочке. Репродукция вируса в культуре клеток сопровождается цитопатическим эффектом и образованием характерных цитоплазматических включений (телец Гварниери), имеющих диагностическое значение.
Резистентность. Вирусы оспы обладают довольно высокой устойчивостью к окружающей среде. На различных предметах при комнатной температуре сохраняют инфекционную активность в течение нескольких недель и месяцев; не чувствительны к эфиру и другим жирорастворителям. При температуре 100ºС вирусы погибают моментально, при 60ºС – в течение 15 мин, при обработке дезинфицирующими средствами (фенол, хлорамин) – в течение нескольких часов. Длительно сохраняются в 50 % растворе глицерина, в лиофилизированном состоянии и при низких температурах.
Восприимчивость животных. Заболевание, сходное по клиническим проявлениям с болезнью человека, можно воспроизвести только у обезьян. Для большинства лабораторных животных вирус оспы малопатогенен.
Эпидемиология. Натуральная оспа известна с глубокой древности. В XVII-XVIII вв. в Европе оспой ежегодно болело около 10 млн человек, из них умирало около 1,5 млн. Оспа являлась также главной причиной слепоты. На основании высокой контагиозности, тяжести течения и значительной летальности натуральная оспа относится к особо опасным карантинным инфекциям.
Источником инфекции является больной человек, который заразен в течение всего периода болезни. Вирус передается воздушно-капельным и воздушно-пылевым путями. Возможен контактно-бытовой механизм передачи – через поврежденные кожные покровы.В начале 20-х годов текущего столетия в результате применения оспенной вакцины удалось ликвидировать натуральную оспу в Европе, Северной Америке, а также в СССР (1936). Отечественные ученые В. М. Жданов, М. А. Морозов и др. обосновали возможность осуществления глобальной ликвидации оспы. В 1958 г. по предложению СССР Всемирная организация здравоохранения приняла резолюцию и разработала программу по ликвидации оспы во всем мире, которая была успешно выполнена благодаря глобальной противооспенной вакцинации людей. В 1977 г. в Сомали был зарегистрирован последний случай оспы в мире. Таким образом, оспа исчезла как нозологическая форма.
Патогенез и клиническая картина. Вирус оспы проникает в организм через слизистую оболочку дыхательных путей и реже через поврежденную кожу. Размножившись в регионарных лимфатических узлах, вирусы попадают в кровь, обусловливая кратковременную первичную вирусемию. Дальнейшее размножение вирусов происходит в лимфоидной ткани (селезенка, лимфатические узлы), сопровождается повторным массивным выходом вирусов в кровь и поражением различных систем организма, а также эпидермиса кожи, так как вирус обладает выраженными дерматотропными свойствами. Инкубационный период составляет 8-18 дней. Заболевание начинается остро, характеризуется высокой температурой тела, головной и поясничной болью, появлением сыпи. Для высыпаний характерна последовательность превращения из макулы (пятна) в папулу (узелок), затем в везикулу (пузырек) и пустулу (гнойничок), которые подсыхают с образованием корок. После отпадения корок на коже остаются рубцы (рябины). По тяжести течения различают тяжелую форму («черная» и сливная оспа) со 100% летальностью, среднюю с летальностью 20-40% и легкую с летальностью 1-2%. К числу легких форм натуральной оспы относится вариолоид – оспы у привитых. Вариолоид характеризуется отсутствием лихорадки, малым количеством оспенных элементов, отсутствием пустул или сыпи вообще.
Иммунитет. У переболевших людей формируется стойкий пожизненный иммунитет, обусловленный выработкой антител, интерферона, а также клеточными факторами иммунитета. Прочный иммунитет возникает также в результате вакцинации.
Лабораторная диагностика. Работа с вирусом натуральной оспы проводится в строго режимных условиях по правилам, предусмотренным для особо опасных инфекций. Материалом для исследования служит содержимое элементов сыпи на коже и слизистых оболочках, отделяемое носоглотки, кровь, в летальных случаях – кусочки пораженной кожи, легкого, селезенки, кровь. Экспресс-диагностика натуральной оспы заключается в обнаружении: а) вирусных частиц под электронным микроскопом; б) телец Гварниери в пораженных клетках; в) вирусного антигена с помощью РИФ, РСК, РПГА, ИФА и других специфических реакций. Выделение вируса осуществляют в куриных эмбрионах или клеточных культурах. Идентификацию вируса, выделенного из куриного эмбриона, проводят с помощью РН (на куриных эмбрионах), РСК или РТГА. Вирус, выделенный на культуре клеток, обладает гемадсорбирующей активностью по отношению к эритроцитам кур, поэтому для его идентификации используют реакцию торможения гемадсорбции и РИФ. Серологическую диагностику осуществляют с помощью РТГА, РСК, РН в куриных эмбрионах и на культурах клеток.
Специфическая профилактика и лечение. Живые оспенные вакцины готовят накожным заражением телят или куриных эмбрионов вирусом вакцины (осповакцины). Повсеместная вакцинация населения привела к ликвидации натуральной оспы на земном шаре и отмене с 1980 г. обязательного оспопрививания. Поэтому оспенные вакцины необходимо использовать только по эпидемическим показаниям с целью экстренной массовой профилактики. Методы введения вакцин – накожно или через рот (таб-летированная форма). После вакцинации формируется прочный иммунитет.
Для лечения натуральной оспы, помимо симптоматической терапии, применяли химиотерапевтический препарат – метисазон.
Не нашли то, что искали? Воспользуйтесь поиском:
источник
Источник