Структурно функціональна організація кори великих півкуль
3.10.Структурно-функціональна організація кори великих півкуль
Великі півкулі головного мозку — парні утворення. Права і ліва півкулі розділені глибокою повздовжньою борозною, в глибині якої міститься мозолисте тіло, утворене нервовими волокнами. Мозолисте тіло з’єднує півкулі мозку. Нервові волокна, що виходять з будь-якої точки однієї півкулі, надходять в симетричну точку другої півкулі, тобто спостерігається парна діяльність великих півкуль. Це явище дістало назву білатеральної симетрії.
Мал. 12. Схема півкуль головного мозку |
Кора великих півкуль головного мозку — це тонкий шар сірої речовини /завтовшки від 1,5 до 3 мм./, яка вкриває весь мозок. Кора є молодшим відділом центральної нервової системи. Водночас вона є вищий інтеграційний центр людини, який здійснює функцію мислення, свідомого сприймання сигналів з навколишнього середовища, абстрактного мислення, пам’яті, інтелекту.
Швидкий розвиток нервових структур при порівняно повільному зростанні місткості черепа спричинює утворення зморшок кори у вигляді закруток і борозен. Це дозволяє в малому об’ємі вмістити велику поверхню, яка у людини займає площу близько 0,25м. Лише одна третина поверхні припадає на закрутки, решта залишається у глибині борозен.
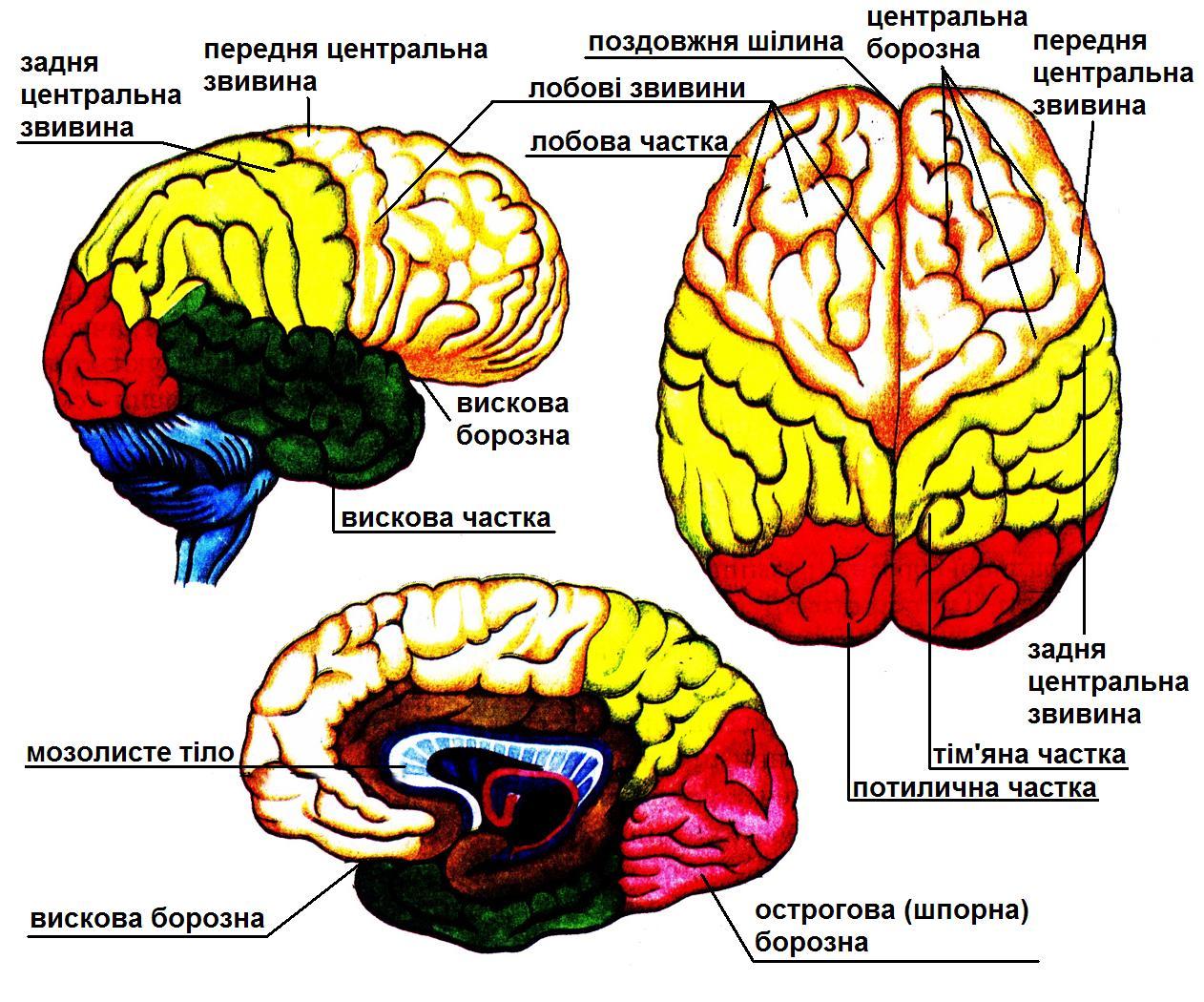
Кора півкуль великого мозку поділяється на чотири частини трьома головними борознами: центральна борозна відділяє лобну частину від тім’яної; тім’яно-потилична борозна — тім’яну від потиличної; бокова борозна відділяє скроневу частину від інших. Кожна частина має ще велику кількість менших борозен, між якими розташовані звивини.
Кора великих півкуль складається з 14-17млрд клітин різних типів: веретеноподібні, зірчасті та пірамідні. Головними є пірамідні і зірчасті нейрони. Зірчасті виконують головним чином аферентну функцію, пірамідні і веретеноподібні — це переважно еферентні нейрони. Для більшості ділянок кори головного мозку характерним є шестишарове розташування нейронів. У різних шарах кори нервові клітини відрізняються формою, величиною і характером розташування.
Відростки нервових клітин кори головного мозку зв’язують різні її відділи між собою або встановлюють контакти кори великих півкуль з розміщеними нижче відділами центральної нервової системи. Відростки нервових клітин, що зв’язують між собою окремі ділянки однієї і тієї ж півкулі, називаються асоціативними. Відростки, що зв’язують симетричні частини обох півкуль, називаються комісуральними, більша частина їх проходить через мозолисте тіло. Проекційні волокна виходять за межі півкуль. Вони входять до складу висхідних і низхідних шляхів, по яких здійснюється двобічний зв’язок кори з відділами центральної нервової системи, що лежать нижче.
Функції кори головного мозку:
1. Кора головного мозку здійснює взаємозв’язок організму з оточуючим середовищем за рахунок безумовних і умовних рефлексів.
2. Вона є основою вищої нервової діяльності / поведінки / організму.
3. За рахунок діяльності кори головного мозку здійснюється вищі психічні функції: мислення і свідомість.
4. Кора головного мозку регулює і поєднує роботу всіх внутрішніх органів, регулює обмін речовин.
Таким чином, з виникненням кори головного мозку вона починає контролювати всі процеси, що протікають в організм, а також всю діяльність людини, тобто відбувається кортиколізація функцій. Причому різні ділянки кори великих півкуль відповідають за певні функції. Такі ділянки кори дістали назву зон. Залежно від ефекту, який відбувається при подразненні, виділяють чутливі / сенсорні/, рухові і неспецифічні зони.
Чутливі зони. Зона шкірної чутливості знаходиться в задній центральній закрутці. Клітини цієї зони сприймають імпульси від тактильних, больових і температурних рецепторів шкіри. Ураження цієї зони на одній стороні призводить до порушення шкірної чутливості на протилежній стороні тіла. При двохсторонньому пошкодженні вказаної зони кори головного мозку спостерігається повна втрата чутливості /анестезія/.
Зорова чутлива зона розташована в потиличній області кори, куди йдуть імпульси від рецепторів сітківки ока. При ураженні цієї зони може порушитись зорова пам’ять, орієнтація в незвичній обстановці і розвинутись повна кіркова сліпота.
Слухова чутлива зона локалізується в скроневих кори головного мозку. Сюди надходять імпульси від рецепторів внутрішнього вуха. При ураженні цієї зони може виникнуть музикальна і мовна глухота, коли людина чує, але не розуміє значення слів. Двостороннє ураження даної області призводить до повної глухоти.
Зона смакової чутливості розміщена в нижніх долях центральної борозни. Ця область отримує нервові імпульси від смакових рецепторів слизової оболонки ротової порожнини. Ураження цієї зони призводить до втрати або потворностей смакових відчуттів.
Зона відчуття запахів розташована в передній частині грушовидної області кори головного мозку. Сюди надходять нервові імпульси від нюхових рецепторів слизової оболонки носа. Ураження цієї зони веде до зниження або втрати нюху.
Таким чином, головна функція чутливих ділянок полягає в критичній оцінці та інтеграції інформації, яка надходить із специфічних ядер зорового горба.
Рухові зони. Ділянки кори, подразнення яких викликає чіткі рухові реакції, називають руховими зонами. Моторна зона локалізується в передній центральній звивині і розташованих поблизу ділянках лобної області. При подразненні передньої центральної борозни виникають різноманітні скорочення скелетних м’язів на протилежній стороні. В залежності від обширності ураження настають паралічі /втрата рухів/ або парези /послаблення рухів/. При ураженні лобних ділянок у людини порушуються складні рухові функції, що забезпечують трудову діяльність і мову, а також пристосовані поведінкові реакції організму.
Рухова і чутливі зони взаємодіють і у функціональному відношенні являють собою єдинеціле, тому їх об’єднують у єдину чутливо-рухову зону.
Неспецифічні ділянки — це ділянки мозку, які не виконують безпосередньо чутливих або рухових функцій. В корі великих півкуль знаходиться декілька зон, що відають функцію мови. В лобній частині лівої півкулі /у праворуких/ розміщений моторний центр мови. При його ураженні мова ускладнюється, або навіть стає неможливою. В скроневій області знаходиться сенсорний центр мови, ушкодження якого призводить до розладів сприйняття мови: хворий не розуміє значення слів, хоча здатність вимовлять їх зберігається. В потиличній ділянці є зони, що забезпечують сприйняття письмової /зорової/ мови. При їх ураженні хворий не розуміє написаного.
Тім’яна область не сприяє інформації ні від яких рецепторів, тому її відносять до асоціативних зон. Клітини цієї області разом з клітинами скроневої ділянки беруть у формуванні мови, сприйнятті форми і розташування власного тіла та просторового співвідношення навколишнього світу.
Лобова частина є нервовою структурою, яка організує реакції поведінки, використовуючи накопичений досвід. Діяльність середніх ділянок лобових часток пов’язана з формуванням особистості і соціальних відносин людей. У хворих з ураженням лобових часток відмічають зміни особистості — зменшуються і навіть повністю зникають мотивації, тверді наміри і плани, які людина прогнозувала. Таким чином, є всі підстави вважать, що лобова частка відіграє головну роль у виробленні стратегії поведінки.
Але будь-яка функціональна зона кори головного мозку знаходиться в аналітичному і функціональному контакті з іншими зонами коривеликих півкуль, з підкірковими ядрами, з утворами проміжного мозку і ретикулярної формації, що забезпечує досконалість виконуючих ними функцій.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Источник
Роль кори головного мозку в системній організації діяльності організму
Роль кори головного мозку в системній організації діяльності організму
План
- Структурно-функціональні особливості кори
- Роль кортикальних колонок
- Проекційні та асоціативні поля
- Методи вивчення кори великих півкуль
- Електроенцефалографія
Кора великих півкуль зявляється вперше у риб у вигляді нюхових клітин, але видалення їх суттєво не впливає на поведінку риб. В амфібій вже є нюхова кора. Півкулі зявляються лише у птахів, у них при видаленні півкуль помітно змінюється поведінка: самі не можуть злетіти, самі не можуть їсти, вють гнізда, але не реагують на крик пташенят, у них зникає материнський інстинкт. Повного розвитку кора досягає у ссавців. У них видалення кори викликає втрату материнського інстинкту (собаки можуть поїдати своїх щенят), тварини не обходять перепони, натикаються на предмети, не йдуть на клич господаря, не орієнтуються в просторі, у них зникає захисний інстинкт. Найбільшу площу займає кора у приматів 2200 см2. В них найкраще розвинені борозни та звивини. Функції кори повністю забезпечують пристосування до життя та вищу психічну діяльність. Видалення великих півкуль призводить до втрати здатності до самостійного життя.
У людини кора великих півкуль забезпечує такі функції:
— Взаємодія організму з навколишнім середовищем.
— Регуляція діяльності внутрішніх органів.
— Регуляція обміну речовин та енергії.
— Вища нервова діяльність мова, память, мислення, свідомість.
Кора головного мозку є вищим відділом ЦНС. Це сіра речовина товщиною 3-5 мм, вкриває півкулі головного мозку. Вона займає площу 22 м2, утворюючи багаточисельні складки. В складі кори до 109-1010 нейронів, які утворюють 6 шарів:
- Молекулярний шар має мало клітин, їх волокна утворюють поверхневе густе тангенціальне сплетіння з дендритами другого шару.
- Зовнішній зернистий шар пірамідні клітини середньої величини, волокна яких розташовані радіально.
- Внутрішній зернистий шар зірчасті клітини, волокна яких розташовані горизонтально.
- Внутрішній пірамідний (гангліозний) шар це гігантські пірамідні клітини Беца, які мають довгі дендрити, що тягнуться до молекулярного шару.
- Поліморфний шар це шар веретеноподібних клітин.
Звязок кори великих півкуль з підкорковими структурами здійснюється за допомогою аферентних і еферентних волокон. Аферентні волокна називаються кортикопетальними, вони несуть інформацію в кору. Основними з них є таламокортикальн волокна. Це прямі аферентні шляхи, які розгалужуються у внутрішньому зернистому шарі і не дають колатералей. Невелика частина волокон йде в молекулярний шар, утворюючи колатеральні еферентні волокна, які називаються кортикофугальними, вони несуть інформацію від кори до підкоркових структур. Ці волокна діляться на 3 групи:
- Проекційні прямі еферентні волокна, що утворюють провідні шляхи (кортикоспінальні, кортикоталамічні)
- Асоціативні — волокна, що утворюють безліч колатералей та йдуть в різні підкоркові зони однойменної півкулі.
- Комісуральні — волокна, що йдуть від кори в складі мозолистого тіла і зєднують зони кори однієї півкулі з підкорковими зонами другої.
1, 2 шари кори великих півкуль забезпечують аналіз та синтез отриманої інформації, мають багато асоціативних волокон.
3, 4 шари кори великих півкуль одержують інформацію від усіх органів та частин тіла за рахунок кортикопетальних волокон.
5, 6 шари кори великих півкуль це рухові нейрони, звідси починаються рухові шляхи, що включають кортикофугальні волокна.
В шарах клітини розміщуються перпендикулярно до поверхні кори, утворюючи ланцюги. Елементарні нервові ланцюги відповідають за переробку певної інформації. Такий функціональний принцип названо кортикальні колонки. Це елементарна функціональна одиниця, в якій здійснюється локальна переробка інформації від рецепторів однієї модальності. Кожна колонка має діаметр 500-1000 мкм, в складі яких розміщується 5-6 нейронів. Пірамідні клітини орієнтовані вертикально, їх аксони утворюють зворотні колатералі, які забезпечують як процеси полегшення в межах мікромодуля, так і гальмування між мікромодулями. Аксони зірчастих клітин ідуть через інтернейрони горизонтально, тому вони, головним чином, забезпечують гальмівні процеси. Веретеноподібні клітини мають довгі аксони, які орієнтовані як горизонтально, так і вертикально. Вони формують кортико-таламічні шляхи.
Мікромодулі обєднуються в макромодулі завдяки горизонтальним розгалуженням терміналей. В колонці можуть бути прості та складні нейрони. Поряд з цим, в корі існує система, яка зчитує елементарні процеси в колонках та обєднує всю інформацію. Формування таких систем зумовлено внутрішньо-кортикальними звязками між окремими макромодулями. Збудження одного мікромодуля викликає гальмування сусідніх. Активація мікромодулів відбувається за рахунок горизонтальних волокон таламокортикальних шляхів.
За функціями поля кори великих півкуль неоднозначні. Вивчення ролі окремих зон кори методом подразнення стало основою для вчення про локалізацію функцій в корі великих півкуль. Пізніше І.П. Павлов методом видалення в поєднанні з методом умовних рефлексів, підтвердив локалізацію функцій в корі. Разом з тим, Лешлі та ін. вважали, що різні зони кори великих півкуль є еквіпотенціальними, тобто однаковими за функціями так виникла теорія еквіпотенціальності. Така думка була помилковою, бо тільки у нижчих тварин кора не має спеціалізованих зон. Так Лешлі, зруйнувавши кору щурів, знайшов, що порушення навиків руху щурів по лабіринту не залежить від місця руйнування. Досліди на тваринах з використанням мікроелектродного методу показали, що зони кори неоднозначні за функціями. Проекційні поля, створені мономодальними нейронами, одержують інформацію від рецепторів через переключаючі ядра таламуса. Вони оцінюють вид подразнення. На їх долю припадає 14% кори. Це специфічні проекційні поля. Асоціативні поля (86%) знаходяться навколо проекційних полів. Це полімодальні нейрони, які одержують інформацію від асоціативних ядер таламуса. Вони оцінюють джерело подразнення, його властивості та відстань до нього. Серед асоціативних полів є вищі поля, які забезпечують психічну діяльність людини, а також є поля, що формують память, індивідуальний досвід, прогнозування, передбачення. Це орбіто-фронтальні поля та тімяна кора. Ці поля співпадають з функціональними зонами, де знаходяться нервові центри.
Представництво всіх видів чутливості в задній центральній звивині.
Задня центральна звивина поля по Бродману 1, 2, 3 корковий центр всіх видів шкірної та пропріоцептивної чутливості. Вона має такі особливості:
- Проекція протилежної сторони тіла розміщена вниз головою.
- Має місце соматотопічна організація для кожної частини тіла.
- Поля шкірної чутливості неоднозначні за площею, так, проекційні поля для обличчя, губ, кистей рук займають найбільшу площу.
При подразненні цієї зони виникає відчуття оніміння, тиску, повзання мурашок. При руйнуванні її зникає чутливість на протилежній стороні анестезія.
Проекція рухових центрів в передній центральній звивині.
Передня центральна звивина поля по Бродману 4, 6 моторна зона. Вона забезпечує згинання, розгинання, пронацію, супінацію, ротацію. Вона також має соматотопічну організацію, але тут найбільшу площу займають мімічні мязи обличчя, язик, кисті рук. При подразнення цієї зони виникає скорочення мязів протилежної сторони тіла. При пошкодженні розвиваються парези та паралічі. Парез це обмеження довільних рухів. Параліч це відсутність рухів.
Сенсорні системи мають подвійну організацію:
а) SS1 це сомато-сенсорна зона І, знаходиться в задній центральній звивині, виконує дискримінативний аналіз, тобто оцінює форму і характер поверхні предмету, приймає участь в складних рухах;
б) SS2 це сомато-сенсорна зона ІІ, має відношення до формування регуляції довільних рухів, орієнтації голови у напрямку звуків. Обидві зони мають сенсорні та моторні входи.
в) SМ1 це сомато-моторна зона І, знаходиться в передній центральній звивині;
г) SМ2 це сомато-моторна зона ІІ, знаходиться там же на медіальній поверхні кори. Обидві зони забезпечують відчуття просторової протяжності, стереогноз, відчуття ваги.
Потилична кора це корковий центр зору. Він оцінює вигляд, форму, розміри та колір предмету. При пошкодження виникає геміанопсія (випадає частина поля зору), або виникає зорова агнозія (людина не впізнає знайомі предмети).
Верхня скронева звивина корковий центр слуху (звивина Гешля поля 41, 42). Оцінює звукові подразнення. При пошкодженні людина не може повторити сказане слово, хоч і чує, людина не розуміє звернену до неї мову, це слухова агнозія.
Задня частина нижньої лобної звивини (центр Брока) це моторний центр мови. У правців він розміщується зліва. При пошкодження людина втрачає здатність до виразного мовлення, узгодження слів у реченні моторна афазія.
Задня частина верхньої лобної звивини (центр Верніке) це сенсорний центр мови, відповідає за сприйняття мови. При пошкодженні людина не розуміє звернену до неї мову сенсорна афазія.
Задній відділ середньої лобної закрутки руховий центр письма. При пошкодженні людина втрачає здатність писати аграфія.
Кутова закрутка нижньої тімя
Источник
Структурно-функціональна характеристика типів кори
Мы поможем в написании ваших работ! Мы поможем в написании ваших работ! Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ? | великих півкуль(по К. Бродману)
Вивчення нейронної організації кори великого мозку дозволило С.Рамон-і-Кахалю, Я.Сентаготаю і А.М. Антоновій висловити думку про модульний характер її будови. Кірковий модуль (нейронний ансамбль) представляє собою групу нейронів, а також гліальних клітин і кровоносних судин, особливим образом розташованих і функціонально зв’язаних між собою. Такий модуль забезпечує обробку і зберігання інформації в корі мозку. Кірковий модуль частіше має вигляд дискретного колонного блоку клітин діаметром 300-600 мкм, який охоплює у вертикальному направленні всі коркові шари. З модулем пов’язаний певний набір аферентних волокон, які приносять інформацію, що підлягає стандартній дискретній обробці, а також набір еферентних волокон, що доставляють цю інформацію в певні зони мозку. Різні модулі кори мозку тісно зв’язані між собою за допомогою інтернейронів і внутрішньокіркових волокон. За допомогою морфологічних і фізіологічних методів отримані численні докази того, що принцип модульної структурно — функціональної організації характерний для всіх відділів центральної нервової системи. БАЗАЛЬНІ ЯДРА Базальні ядра(nuclei basales) представляють собою скупчення сірої речовини в товщі білої речовини великих півкуль. У сірій речовині розрізняють смугасте тіло, огорожу і мигдалевидне тіло. Смугасте тіло (corpus stratum) складається з двох утворень: хвостатого ядра і сочевицеподібного ядра. Сіра речовина хвостатого і сочевицеподібного ядер розташована між прошарками білої речовини, що і обумовило загальну назву цієї групи ядер — смугасте тіло. Хвостате ядро представляє собою округле вигнуте і витягнуте тіло, яке розташоване вище сочевицеподібного ядра (мал. 53). Передня розширена частина хвостатого ядра (головка) утворює бічну стінку переднього рогу бічного шлуночка мозку. Головка спереду прилягає до передньої продірявленої речовини і з’єднується із лушпиною. Задня вигнута і загострена частина (хвіст) формує частину дна середньої частини бічного шлуночка і продовжується до нижнього рогу бічного шлуночка, закінчуючись поблизу мигдалевидного тіла. Медіальна поверхня хвоста прилягає до верхньо-бічної поверхні таламуса. Мал. 53. Горизонтальний розріз головного мозку на рівні базальних ядер півкуль: 1-коліно мозолистого тіла, 2-передній ріг бічного шлуночка, 3-прозора перегородка, 4-ніжки склепіння, 5-сочевицеподібне ядро, 6-внутрішня капсула, 7-таламус, 8-мозолисте тіло, 9-черв’як мозочка, 10-задній ріг бічного шлуночка, 11-порожнина третього шлуночка. 12-зовнішня капсула, 13-огорожа, 14-хвостате ядро (мал. В.І. Козлова). Сочевицеподібне ядро розташоване попереду від таламуса і відокремлено від нього білою речовиною — внутрішньою капсулою, в якій проходять шляхи, що з’єднують кору великих півкуль з нижче розташованими відділами мозку. Сочевицеподібне ядро складається з більш темної латеральної частини — лушпини і більш світлої медіальної частини — блідої кулі. Обидві частини розділені між собою смушками білої речовини. Лушпина з’єднується з хвостатим ядром тонкими тяжами сірої речовини, що розміщуються між пучками волокон внутрішньої капсули. Хвостате ядро і лушпина мають філогенетично більш пізнє походження, порівняно з блідою кулею. Обидві утворення мають схожу нейронну структуру і їх об’єднують під загальною назвою стріатум, або неостріатум. Стратум і бліда куля (палідум) пов’язані з корою великих півкуль, таламусом, ядрами стовбура мозку, мозочком. Завдяки цьому, стріатум і палідум, а також їх зв’язки складають стріо-палідарну систему. Ця система управляє складнокоординованими автоматизованими рухами організму, контролює і підтримує тонус скелетних м’язів, а також є вищим центром регуляції таких вегетативних функцій як, теплопродукція і вуглеводний обмін у мускулатурі. При пошкодженні шкарлупи і блідої кулі можуть спостерігатись повільні стереотипові рухи (атетоз). Із стріатумом і палідумом функціонально і анатомічно тісно зв’язана чорна речовина середнього мозку. При дегенерації шляхів від чорної субстанції до стріатуму розвивається паркінсонізм з характерним сильним тремором (тремтіння) кінцівок і голови та труднощами в управлінні рухами внаслідок порушення регуляції м’язового тонусу з боку стріо-палідарної системи. Огорожа — це вертикальна пластинка сірої речовини, нижня частина якої продовжується в речовину передньої продірявленої пластинки на основі мозку. Огорожа розташована з боку сочевицеподібного ядра і відокремлена від нього зовнішньою капсулою мозку. Від кори закруток острівця огорожу відокремлює тонкий шар білої речовини зовнішньої капсули мозку. Огорожа є найбільш молодим у філогенетичному відношенні утворенням серед базальних ядер і має численні зв’язки з корою великих півкуль. Мигдалевидне тіло- знаходиться в ділянці полюса вискової частки і межує з гіпокампом. До його складу входять три групи ядер: бічна, яка зв’язана з корою великих півкуль, кірково-присередня — зв’язана із структурами нюхової системи, і центральна — зв’язана з гіпоталамусом і ядрами стовбуру мозку, яка контролює вегетативні функції організму. Мигдалевидне тіло є важливою частиною лімбічної системи мозку. Його руйнування приводить до агресивної поведінки або апатичному, млявому стану. Завдяки своїм зв’язкам з гіпоталамусом мигдалевидне тіло впливає на ендокринну систему, а також на репродуктивну поведінку. Базальні ядра відмежовуються одне від одного прошарками білої речовини: внутрішньою і зовнішніми капсулами (мал. 53). Внутрішня капсула розташована між хвостатим ядром і таламусом з однієї сторони, і сочевицеподібним ядром — з іншої. Вона складається з проекційних волокон висхідних і низхідних шляхів, які з’єднують кору головного мозку із стовбуром мозку і спинним мозком. Волокна, що проходять ростральніше утворюють передню ніжку внутрішньої капсули, а каудальніше — задню ніжку. У передній ніжці проходять кірково — таламічні волокна, а також волокна від лобової частки до власних ядер моста і далі до мозочка. У задній ніжці волокна ідуть до кори великого мозку від органів чуття і таламуса. Тут також ідуть низхідні волокна у складі кірково-спинномозкових шляхів, а також від тім’яної, вискової і потиличної часток до власних ядер моста. Зовнішні капсули відмежовують огорожу від сочевицеподібного ядра і від кори острівцевої частки. В цих капсулах проходить велика кількість волокон, які з’єднують огорожу з різними зонами кори великих півкуль. МОЗКОВІ ОБОЛОНКИ І КРОВОНОСНІ СУДИНИ ГОЛОВНОГО МОЗКУ Головний мозок оточений трьома оболонками: внутрішньою, або м’якою, середньою, або павутинною, і зовнішньою, або твердою. Всі вони є продовженнями оболонок спинного мозку. М’яка (або судинна) оболонкаприлягає до мозку і, повторюючи його рельєф, заходить в усі борозни. Вона містить кровоносні судини і утворює судинні сплетення, які розташовані в шлуночках мозку. Судинні сплетення продукують спинномозкову рідину, яка циркулюючи у мозкових шлуночках і в підпавутинному просторі головного і спинного мозку, захищає їх від механічних впливів і виконує роль лімфи. Судинні сплетення також затримують і знешкоджують шкодливі речовини. Павутинна оболонка у борозни не заходить, а, перекидаючись між закрутками мозку, утворює особливі підпавутинні простори — цистерни, в яких циркулює спинномозкова рідина. Найбільш великими цистернами є: цистерна бічної ямки великого мозку, міжніжкова і мозочково-мозкова цистерни. Підпавутинні простори спілкуються між собою, з шлуночками мозку і з підпавутинними просторами спинного мозку. Тверда оболонка (мал. 54) дає у порожнину черепа вирости, які розташовані між окремими частинами головного мозку і захищають його разом із спинномозковою рідиною від струсу. Найбільш важливі з цих відростів: серп великого мозку, який проникає в повздовжню борозну між півкулями, і намет мозочка, що відокремлює півкулі головного мозку від мозочка. Крім цього, тверда мозкова оболонка в порожнині черепа утворює канали — синуси, по яких здійснюється відтік венозної крові з мозку. Основними синусами твердої мозкової оболонки є: верхній і нижній сагітальні, лівий і правий поперечні, лівий і правий сигмоподібні. Із синусів венозна кров відтікає через яремні отвори черепа у внутрішні яремні вени. Кровопостачання головного мозку здійснюється з двох джерел: внутрішніх сонних артерій і хребетних артерій (мал. 55). Ці артерії утворюють на основі головного мозку артеріальне коло великого мозку. Внутрішня сонна артерія, після заходження в порожнину черепа, розгалужується на передню і середню мозкові артерії. Передня мозкова артерія дає кров, головним чином, до лобової частки мозку, середня мозкова артерія — до тім’яної і вискової часток, а очна артерія (яка є продовженням передньої мозкової артерії) — до очного яблука. Передні мозкові артерії з’єднуються поперечним анастомозом — передньою сполучною артерією. Хребетні артерії (права і ліва) на основі черепа з’єднуються і утворюють непарну базилярну артерію, яка дає гілки до мозочка та інших відділів стовбуру мозку, і дві задні мозкові артерії, які дають кров до потиличної частки мозку. Кожна із задніх мозкових артерій з’єднується із середньою мозковою артерією своєї сторони за допомогою задньої сполучної артерії. Таким чином, на основі мозку утворюється артеріальне коло великого мозку, яке забезпечує найбільш сприятливі умови для функціонування цього життєво важливого органу. Мал. 54. Тверда мозкова оболонка і венозні синуси: 1-серп великого мозку, 2,7-верхній сагітальний синус, 3-прямий синус, 4.6-поперечний синус, 5-намет мозочка, 8-нижній сагітальний синус (мал. В.І. Козлова). Мал. 55. Артерії основи головного мозку: 1-передня сполучна а., 2,13-внутрішня сонна а., 3,12-задня сполучна а., 4,11 — задня мозкова а., 5-передня нижня мозочкова а., 6,9-хребетна а., 7-задня нижня мозочкова а., 8-передня спинномозкова а., 10-базилярна а., 14-середня мозкова а., 15-передня мозкова а.. (мал. В.І. Козлова). ПРОВІДНІ ШЛЯХИ ЦЕНТРАЛЬНОЇ НЕРВОВОЇ СИСТЕМИ Провідні шляхи ЦНС побудовані із функціонально однорідних груп нервових волокон. Вони представляють собою внутрішні зв’язки між ядрами і кірковими центрами, розташованими в різних частинах і відділах мозку, і служать для їх функціонального об’єднання (інтеграції). Провідні шляхи, як правило, проходять у білій речовині спинного і головного мозку, але можуть локалізуватись і в покришці стовбуру мозку, де чіткої межі між білою і сірою речовинами немає. Основною провідною ланкою в системі передачі інформації від одних центрів до інших є нервові волокна — аксони нейронів, які передають інформацію у формі нервового імпульсу в строго визначеному напрямку, а саме від тіла клітини. Серед провідних шляхів в залежності від їх будови і функціонального значення виділяють різні групи нервових волокон: волокна, пучки, тракти, спайки (комісури), променистості. ВНУТРІШНІ ЗВ’ЯЗКИ ГОЛОВНОГО І СПИННОГО МОЗКУ Серед внутрішніх зв’язків у ЦНС розрізняють три групи провідних шляхів: асоціативні, комісуральні і проекційні. Асоціативні шляхи досить численні і широко представлені в різних відділах ЦНС, але найбільш розвинуті вони в корі мозку. Ці шляхи утворені асоціативними нейронами та їх волокнами, які з’єднують різні ділянки кори мозку однієї півкулі або мозочка. Волокна можуть проходити усередину сірої речовини, утворюючи внутрішньокіркові горизонтальні пучки і знаходитись у складі білої речовини мозку. Асоціативні шляхи з’єднують також різні ділянки однієї половини спинного мозку (табл. 3). Асоціативні волокна в півкулі мозку можуть бути короткими і довгими. Короткі волокна (дугоподібні волокна) з’єднують між собою сусідні закрутки і включають в основному аксони нейронів VI і VII цитоархітектонічних шарів кори. Довгі волокна є аксонами пірамідних нейронів переважно ІІІ і V кіркових шарів і утворюють пучки, які з’єднують між собою віддалені один від одного ділянки кори або частки великого мозку. Серед асоціативних пучків великих півкуль розрізняють пояс, який з’єднує між собою різні ділянки лімбічної частки. До них належать також верхній повздовжній пучок, що з’єднує лобову частку з нижньою тім’яною часточкою, потиличною часткою і задньою частиною вискової частки. Нижній повздовжній пучок з’єднує потиличну і вискову частки, а гакоподібний пучок — нижню поверхню лобової частки і висковий полюс. Таблиця 3 |
Источник