Слухова зона кори головного мозку
Содержание статьи
луховая кора. Слуховая функция коры головного мозга
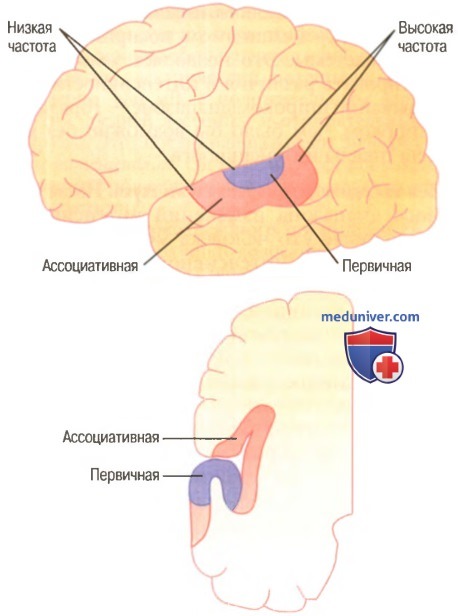
Слуховая кора. Слуховая функция коры головного мозгаНа рисунке ниже показана проекционная слуховая зона мозговой коры, которая лежит главным образом в надвисочной плоскости верхней височной доли, но также распространяется на латеральную сторону височной доли, на большую часть островковой коры и даже на латеральную часть теменной покрышки.
На рисунке выше показаны две отдельные части слуховой коры: первичная слуховая кора и ассоциативная слуховая кора (называемая также вторичной слуховой корой). Первичная слуховая кора непосредственно возбуждается проекциями от медиального коленчатого тела, тогда как ассоциативная слуховая кора возбуждается вторично импульсами из первичной слуховой коры, а также проекциями из таламических ассоциативных зон, прилежащих к медиальному коленчатому телу. а) Восприятие частоты звука в первичной слуховой коре. В первичной слуховой коре и ассоциативной слуховой коре обнаружены, по крайней мере, шесть тонотопических карт. В каждой из этих карт высокочастотные звуки возбуждают нейроны у одного конца карты, а низкочастотные звуки — у противоположного конца. В основном, как видно на рисунке выше, низкочастотные звуки локализуются спереди, а высокочастотные — сзади. Это справедливо не для всех карт. Возникает вопрос: почему слуховая кора имеет так много разных карт? Ответ, вероятно, в том, что каждая из отдельных областей анализирует одно из специфических свойств звука. Например, одна из больших карт в первичной слуховой коре, вероятно, различает сами звуковые частоты и дает человеку физическое ощущение высоты звуков. Другая карта, вероятно, используется для определения направления, откуда исходит звук. Другие области слуховой коры выделяют особые качества, например внезапное начало звуков, или, возможно, особые модуляции, например выделение звуков определенной частоты из шума. Диапазон частот, на которые реагирует каждый нейрон слуховой коры, гораздо уже, чем в улитке и релейных ядрах мозгового ствола. Вновь обратившись к рисунку, можно видеть, что базальная мембрана улитки стимулируется звуками всех частот, и такое же широкое звуковое представительство обнаруживается в улитковых ядрах. Однако когда возбуждение достигает мозговой коры, большинство звукочувствительных нейронов реагируют лишь на узкий, а не на широкий диапазон частот. Следовательно, где-то по пути механизмы анализа «обостряют» реакцию на частоту. Полагают, что причиной этого обостряющего эффекта является главным образом феномен латерального торможения. Это значит, что стимуляция улитки одной частотой тормозит звуковые частоты с обеих сторон этой первичной частоты; причиной являются коллатеральные волокна, ответвляющиеся от первичного сигнального пути и оказывающие тормозное влияние на прилежащие пути. Важность такого эффекта показана также для усиления особенностей соматосенсорных, зрительных и других типов ощущений. Многие нейроны слуховой коры, особенно в ассоциативной слуховой коре, не просто реагируют на специфические звуковые частоты в ухе. Полагают, что эти нейроны «связывают» разные звуковые частоты друг с другом или звуковую информацию с информацией от других сенсорных областей коры. Действительно, теменная часть ассоциативной слуховой коры частично перекрывает соматосенсорную область II, что, вероятно, обеспечивает возможность ассоциации слуховой информации с соматосенсорной. — Также рекомендуем «Различение звуковых образов. Определение направления звука» Оглавление темы «Физиология слуховой системы»: |
Источник
Зони кори великого мозку
Окремі ділянки кори мають різне функціональне значення. Разом з підкірковими центрами, стовбуром мозку і спинним мозком великий мозок об’єднує окремі частини організму в єдине ціле, здійснює нервову регуляцію всіх органів (рис. 55, 56).
У кору великого мозку надходять доцентрові імпульси від рецепторів. Кожному рецепторному апарату відповідає в корі ділянка, яку І.П Павлов назвав кірковим ядром аналізатора. Ділянка кори, де розташовані кіркові ядра аналізаторів, названі сенсорними зонами кори великого мозку.
Ядерна зона рухового аналізатора (сомато-сенсорна зона), куди надходять збудження від рецепторів суглобів, скелетних м’язів і сухожилок, розташована в передню- і задньоцентральних ділянках кори. У межах передньої центральної закрутки найвище розміщені центри для м’язів нижньої кінцівки, нижче — для м’язів тулуба, потім верхньої кінцівки і, нарешті, центри м’язів голови. Ураження цієї зони призводить до паралічу протилежної половини тіла.
Рис. 55. Зовнішня будова головного мозку (за дорлінг кіндерслі, 2003)
Передцентральна звивина (gyrus precentralis) і прицентральна часіка (lobulus paracentralis) лобової частки становлять руховий центр кори і є аналізатором кінестезичних імпульсів, які надходять від посмугованих м’язів, суглобів, сухожилків. Тут замикаються рухові умовні рефлекси. У верхній ділянці перед центральної звивини розташовані клітинні групи, що належать до м’язів нижніх кінцівок, нижче — верхніх кінцівок, ще нижче — неврони, пов’язані з іннервацією м’язів голови. Оскільки нервові шляхи перехрещуються, праві рухові центри кори пов’язані з мускулатурою лівої сторони тіла і навпаки.
У задній частині середньої лобової звивини міститься центр узгодженого руху голови й очей (окоруховий, блоковий, відвідний і додатковий нерви).
Рис. 56. Кіркові поля (за Дорлінг Кіндерслі, 2003)
У задньому відділі нижньої лобової звивини розміщена зона Брока — руховий центр мови, який разом із центром Верніке забезпечують здатність людини читати, писати, чути, вимовляти і розуміти мову.
Ушкодження різних полів кори лобової частки може призвести: до підвищення агресивності й послаблення реакції страху; зростання пасивно-захисних умовних рефлексів; порушення харчових і захисних умовних рефлексів.
У таких людей спостерігається втрата ініціативи, апатія, порушення абстрактного мислення, нездатність до творчого мислення, розгальмування нижчих емоцій і потягів, розлади мовлення і понятійного мислення.
У задній частині лобової звивини розташований центр письма, ураження якого призводить до порушення навичок письма під контролем зору.
У лівій (у лівшів у правій) нижній тім’яній часточці розташований центр, який координує цілеспрямовані рухи. Він функціонує за типом тимчасових зв’язків, які виникають протягом індивідуального життя, тобто умовних рефлексів. У разі ушкодження цього центру елементи довільних рухів зберігаються, але порушуються цілеспрямовані дії (апраксія).
У верхній тім’яній частці (задньоцентральна звивина) розміщений кірковий центр аналізаторів чутливості (больової, температурної, дотикової), або сомато-сенсорна кора. Ураження кори у цій частині призводить до часткової або повної анестезії (втрата чутливості).
Ураження кори в ділянці верхньої тім’яної частки призводить до зниження больової чутливості і порушення стереогноза — впізнавання предметів на дотик без допомоги зору.
У нижній тім’яній частині розташований центр праксії, який регулює здатність здійснювати координаційні рухи, які складають основу робочих процесів і потребують спеціального навчання.
У кутовій звивині тім’яної частки розташований зоровий центр мови. Його ураження призводить до неможливості розуміння письма (алексія).
Тім’яна ділянка — це апарат вищої інтегративної діяльності мозку людини, вона безпосередньо стосується процесів біологічної і соціальної адаптації, є фізіологічною основою вищих психічних функцій.
Локалізація статичного аналізатора (центр збереження рівноваги і положення тіла в просторі) — кора верхньої та середньої скроневих звивин. Ушкодження цього центру призводить до атаксії (розладу координації рухів).
Зона шкірного аналізатора, зв’язаного з температурою, больовою і тактильною чутливістю займає задньоцентральну ділянку. Центри чутливості нижчих частин тіла розміщені у верхніх частинах тіла — у нижніх її ділянках.
Найбільшу площу займає кіркове представництво рецепторів кисті рук, голосового аналізатора і обличчя, найменшу — тулуба, стегна і гомілки.
Ядерна зона зорового аналізатора розташована на внутрішній поверхні потиличної ділянки, в зоні шпорної борозни. Ураження цього центру призводить до сліпоти. При порушеннях у сусідніх із шпорною борозною частин кори в ділянці потиличного полюса на медіальній і латеральній поверхнях частки може спостерігатися втрата зорової пам’яті, здатності орієнтації у незнайомій обстановці, порушення функції, пов’язаної із бінокулярним зором (здатності за допомогою зору оцінювати форму предметів, відстань до них тощо).
У корі верхньої скроневої звивини розташована частина слухового аналізатора, а поблизу від бокової борозни — ядерна зона смакового аналізатора. Двостороннє ураження до повної кіркової глухоти.
Нюхова зона розміщена на внутрішній поверхні скроневих ділянок кори. В ділянці середньої і нижньої скроневих звивин розташоване кіркове представництво вестибулярного аналізатора. Ураження цієї ділянки призводить до порушення рівноваги під час стояння і зниження чутливості.
Із сенсорними зонами взаємодіє моторна зона кори великого мозку. Ядерні зони аналізаторів — це ділянки кори, в яких закінчується основна маса провідних шляхів аналізаторів. За межами ядерних зон розташовані розсіяні елементи, куди надходять імпульси від тих же рецепторів, що і в ядро аналізатора.
Центр мови міститься у лівій півкулі. Розрізняють 2 центри мови: руховий (зона Брока), який міститься у нижній частині лобової ділянки і слуховий (зона Верніке), який знаходиться у скроневій ділянці, під заднім кінцем сільвієвої борозни. Центри мови є лише у людини. Мовлення, мислення, почуття і вправні рухи контролюються нейронами, які розміщені в лобовій ділянці головного мозку. Розпізнавання тонів і звуків відбувається в скроневій ділянці. Ця ділянка також бере участь у запам’ятовуванні інформації. Різноманітні сенсорні відчуття, такі як біль, температура усвідомлюються та інтерпретуються в тім’яній ділянці. Потилична ділянка фіксує та інтерпретує зорові образи.
Источник
натомия слуховой коры
Оглавление темы «Физиология проводящего пути слуха.»:
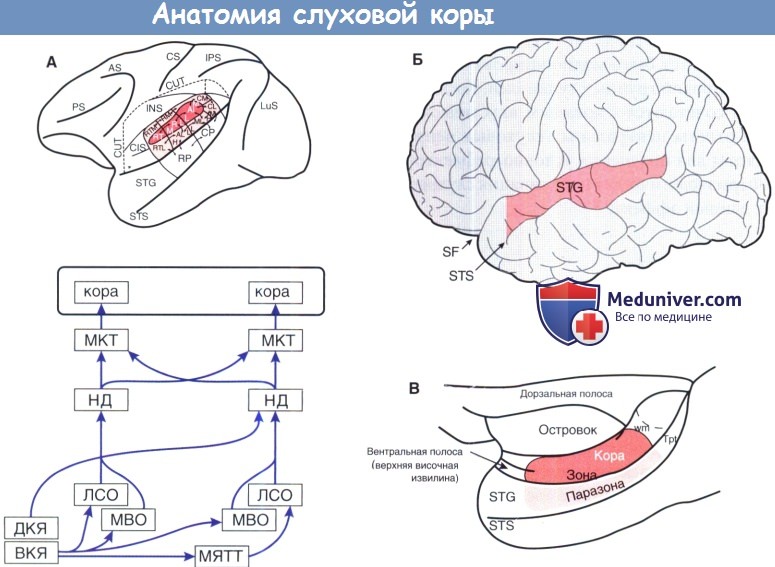
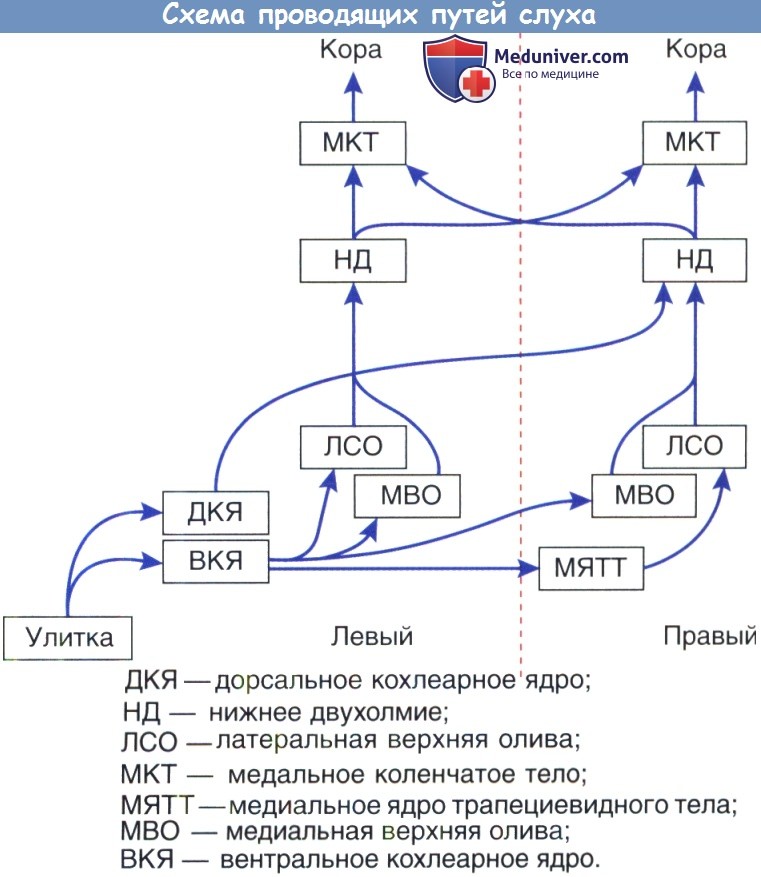
Анатомия слуховой корыСлуховая кора получает все входящие звуковые представления от таламуса. В то время как отдельные подкорковые структуры отвечают за извлечение специфических особенностей из акустических стимулов, слуховая кора должна рекомбинировать эти параллельные потоки информации в интегрированные звуковые восприятия. Эти разнообразные восприятия включают в себя распознавание источников звука, локализацию источника звука, распознавание голосов, интерпретации звуков в биологическом контексте, слуховую память и достигаемую тренировками пластичность. Большая часть наших знаний, относящихся к структуре и функции слуховой коры, была получена в исследованиях на различных видах животных, при этом некоторые из самых интригующих находок получены в работах с приматами. Учитывая ограниченность использования экспериментальных животных в качестве системной модели для изучения высших когнитивных процессов, наши знания о функциях человека значительно продвинулись благодаря косвенным измерениям нервной деятельности, которые могут быть применены к людям, в частности, функциональной нейровизуализации. Технические трудности изучения слуховой функции человека в естественных условиях осложняются выраженной разнохарактерностью слуховой коры с ее многочисленными регионами и полями, каждое из которых обладает своими собственными отличительными анатомическими и физиологическими свойствами. Было использовано много различных анатомических подходов для определения подразделов слуховой коры, в том числе цитоархитектонические и хемоархитектонические методы, в дополнение к изучению нервных связей, участвующих в таламокортикальных, кортико-кортикальных и кортико-фугальных связях. Также были использованы физиологические исследования, основанные на свойствах рецепторного поля, определения НЧ (или частотной характеристики), спектральной полосы целевых нейронов с учетом качества временного ответа. В результате этих исследований четко установлено, что первичное слуховое поле, часто упоминающееся как А1, существует почти во всех видах, включая человека. За А1, однако, было трудно установить единые анатомо-функциональные критерии, которые действительны для разных видов или даже разных индивидуумов одного и того же вида. Анатомия слуховой коры. А1 составляет первую стадию корковой обработки звука. Большая часть того, что нам известно об анатомии А1, основано на исследовании приматов, особенно обезьян, и поэтому, на данный момент, некоторые невыясненные детали у людей могут отличаться. На гистологическом уровне у А1 есть несколько характерных особенностей кониокортекса, включая плотно упакованные маленькие клетки в четвертом слое, обильно миелинизированные волокна и наличие цитохромоксидазы. Как указано ранее, А1 получает большую часть входящих звуков непосредственно от вентральной части таламуса. Функционально, нейроны А1 имеют тенденцию отвечать на чистые тоны хорошо с хорошо настроенными диапазонами НЧ и тонотопическими проведением изочастотного диапазона. Hackett и Kaas используют термин ядро для описания первичных или первично-подобных регионов, которые включают в себя три области (А1, ростральные или Р, и ростротемпоральные или РТ). Эти области представляются активируемыми параллельно, и каждая из этих основных зон демонстрирует отличительные нейронные соединения с другими участками коры. Эти три области организованы каудо-рострально по плоскости боковой щели и имеют тонотопическое расположение. В организме человека большая часть извилины Гешля (поперечная височная извилина) составляет слуховое ядро. Цитоархитектоническим отличием области зоны слуховой коры является то, что она окружает централизованное ядро. Эта область получает несколько входов от вентрального таламического ядра (ВТЯ), его основные таламические входы происходят в дорсальном (ДТЯ), и медиальном ядрах (МТЯ). Нейроны в области зоны получают почти все свои слуховые входы из основных областей и не показывают предварительную активность. Третья область паразоны, точно вентрально к зоне, получает плотные связи от зоны, но почти ни одной от ядра. Паразона не получает таламические входы от дорсального таламического ядра (ДТЯ) и медиального таламического ядра (МТЯ), и корковые входы неслуховых областей прилежат к верхней височной борозде. Эта схема может играть важную роль в полисенсорной обработке (аудиовизуальные взаимодействия). Дополнительные префронтальные связи играют определенную роль в слуховой памяти и зрительно-пространственной ориентации.
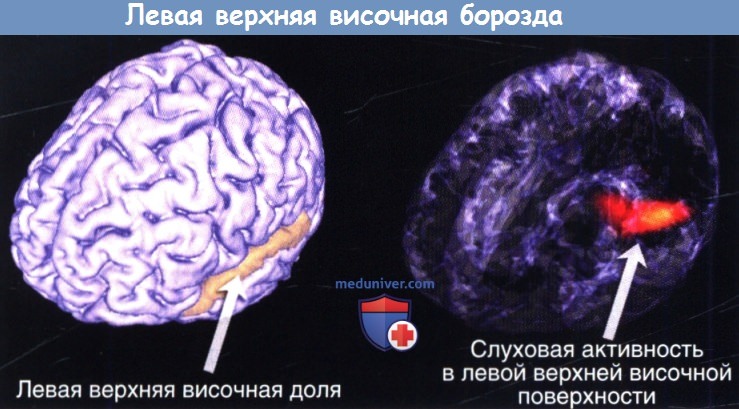
Из областей паразоны и зоны звуковые сигналы поступают на четвертый уровень невральной обработки в пределах височных, париетальных, и лобных долей. Смысл этих отношений заключается в том, что многие участки мозга, даже те, что строго не считаются центрами обработки звуков, получают звуковые импульсы и имеют решающее значение для их правильной обработки. Полисенсорные взаимодействия здесь имеют решающее значение, и необходимы дальнейшие исследования для выяснения точной причины нервных взаимодействий между слуховыми и зрительными участками коры. Кроме того, роль гомологичных областей мозга, связанных с мозолистым телом, очень плохо изучена. Несмотря на это, передача звуковой информации происходит именно восходящим образом, что в конечном итоге позволяет людям осознанное восприятие звукового сигнала. Хотя описания слухового ядра, областей зоны и паразоны у обезьян достаточно понятны, наше понимание человеческой слуховой системы остается относительно неполным, принимая во внимание, что большая часть современных знаний получена из недавних исследований с функциональной визуализацией. В частности, это ограниченное понимание может вызывать сложности при интерпретации результатов исследований поражения слуха у людей. Четко установлено, что слуховая кора у людей концентрируется вдоль верхней височной извилины. Верхняя височная плоскость, включающая плоскость полюса височной извилины, извилину Гешля, височную поверхность (спереди назад) — по-видимому, и есть местоположение слуховой коры человека. Эта область имеет особенно высокую вариабельность от человека к человеку, и разделение коры на поля Бродмана может быть затруднено. В 41-м поле по Бродману, что соответствует передней поперечной височной области (где находится зона А1), по-видимому, присутствует вертикальная столбчатая организация слуховых нейронов (именуемые Tel. 1, Tel.О. и Те1.2 медиальнее или кзади к боковым). Как уточняется в первой половине этой статьи, значительный объем обработки слуховой информации происходит в стволе мозга. Таким образом, слуховая кора получает «предварительные» сигналы, которые затем обрабатываются дальше в ядре, регионах зоны и паразоны. А1, по-видимому, участвует в идентификации звуковых объектов, а также интеграции спектральных и временных свойств звуков. В целом наше понимание не-А1 областей является более скудным по сравнению с А1, в частности, из-за того, что в большинстве исследований приматов использовались электрофизиологические методы, в то время как при исследованиях у людей применялись функциональные методы нейровизуализации. Последний подход имеет низкое временное разрешение и усложняется присутствием сильного фонового «шума» сканера. Например, даже установление точных параметров человеческой тонотопической оси было проблемой, так как не-А1 слабо отвечает на чистые тоны, в то время как А1 отвечает кратковременно. Исследования, где была использована полосовая фильтрация стимулов со спектральными изменениями, убедительно показывают, что человеческая слуховая кора сохраняет тонотопическую ось. Корковая обработка височной информации зависит от скорости передачи информации. В общем, звуки могут быть подразделены на височные структуры замедленного темпа (<30 Гц) и ускоренного темпа (>50 Гц). По замедленному типу височная информация закодирована непосредственно невральными темпами выброса. И извилина Гешля, и не-А1 являются критическими элементами для модуляций замедленного уровня, неотъемлемой частью такого процесса как амплитудное детектирование речи. При ускоренном темпе, опережающем способности частоты невральной реакции, должны использоваться другие стратегии. Височные структуры высокой интенсивности отражаются в диапазоне миллисекунд, что играет роль для результатов исследований РПС (важный для локализации звука) и важно для восприятия музыкальных мелодий.
Несколько исследований показали, что мозг отвечает на височное вариации ускоренного темпа. Невральные реакции на постоянный шум выражены сильнее, чем на контрольный шум, так как высокочастотные колебания вызывают определенные реакции в А1. Только к середине XIX века удалось достигнуть согласия в том, что сенсорно-двигательные функции могут быть локализованы в коре головного мозга. Неприятие функциональной локализации отчасти было связано с реакцией на вводящие в заблуждение теории френологов, которые ограничивали связь сенсорно-двигательных функций только формой головы. В 1861 году при аутопсии пациента с экспрессивной афазией было обнаружено, что левая нижняя лобная извилина была поражена в области, которую с тех пор называют полем Брока. Повреждение задней верхней височной извилины в области, обычно называемой областью Вернике, приводит к сенсорному дефициту, который сразу проявляется клинически. Понятие тонотопии было определено в середине 1900-х, а многие предшествующие исследования специфической роли слуховой коры для частотной избирательности были неубедительны. В 1975 году было обнаружено, что двусторонние слуховые поражения коры приводят к потере способности локализовать источник звука. При удалении у обезьян части коры головного мозга, было определено, что двустороннее удаление верхней височной доли приводит к немедленной невосприимчивости звуковой стимуляции с отсроченным восстановлением в течение нескольких месяцев, в то время как одностороннее удаление приводило к контралатеральной недостаточности. Как было описано ранее, специфические особенности слухового поведения и невральной организации препятствуют нашему пониманию слуховой коры. Критические вопросы, касающиеся и нормальных, и патологических механизмов слуха остаются нерешенными. Конечной целью нейробиологии является понимание того, как работает головной мозг. По отношению к слуховой системе важно понимание того, как центральная и периферические слуховые системы обеспечивают слуховое распознавание и точное восприятие слуховых стимулов. До некоторой степени современное понимание было сформировано подходами, разделяющими центральную нервную систему на меньшие подгруппы и исследующими анатомию и физиология каждой подгруппы отдельно. Следовательно, мы знаем намного больше о каждой отдельной подгруппе, чем об их взаимодействии друг с другом. Кроме того, относительно мало известно, как именно подгруппы слуховых систем реагируют с другими сенсорными системами, чтобы обеспечить слуховое восприятие, которое будет правильно интерпретировано только в рамках реального контекста. Безусловно, полезно подразделить исследование слуховой системы на составляющие части, такие как А1 и вторичная слуховая кора, но у этого подхода есть внутренние ограничения, которые никогда не смогут привести к созданию удовлетворительной модели прослушивания для всего мозга. Представление о специфической «локализации» слуховой функции в отдельной нейроанатомической области, как кратко упоминалось выше, скорее всего слишком упрощенное. Целесообразнее определить функцию каждой подгруппы по порядку, чтобы получить возможность для интерпретации их функционирования в органическом контексте. Поэтому необходимо использование широкого спектра методов и развитие таких стратегий топографии распределения биопотенциалов как функциональная магнитно-резонансная томография (фМРТ), что даст важную информацию о взаимодействии слуховых областей друг с другом, а, возможно, и важные сведения об участии неслуховых областей в поведенческом контексте слушания. Одним из перимуществ фМРТ является то, что информация из разнообразных областей мозга может быть накоплена одновременно, позволяя фиксировать активность в тех областях мозга, связь которых со слуховым восприятием не предполагалась. Важно признать, что многие классические «неслуховые» области могут играть решающую роль при восприятии или генерации сложных звуков.
— Также рекомендуем «Функция и физиология слуховой коры» |
Источник