Рибосомы у вирусов оспы
Содержание статьи
ирусы. Вирион. Морфология вирусов. Размеры вирусов. Нуклеиновые кислоты вирусов.
Оглавление темы «Типы микроорганизмов. Вирусы. Вирион.»: Вирусы. Вирион. Морфология вирусов. Размеры вирусов. Нуклеиновые кислоты вирусов.Вирусы [от лат. virus, яд] — наименьшие по размерам агенты, имеющие геном, окружённый белковой оболочкой. Вирусы не воспроизводятся самостоятельно, они — облигатные внутриклеточные паразиты, репродуцирующиеся только в живых клетках. Все вирусы существуют в двух формах. В настоящее время известны вирусы бактерий (бактериофаги), грибов, растений и животных. Внеклеточная форма — вирион — включает в себя все составные элементы (капсид, нуклеиновую кислоту, структурные белки, ферменты и др.). Внутриклеточная форма — вирус — может быть представлена лишь одной молекулой нуклеиновой кислоты, так как, попадая в клетку, вирион распадается на составные элементы.
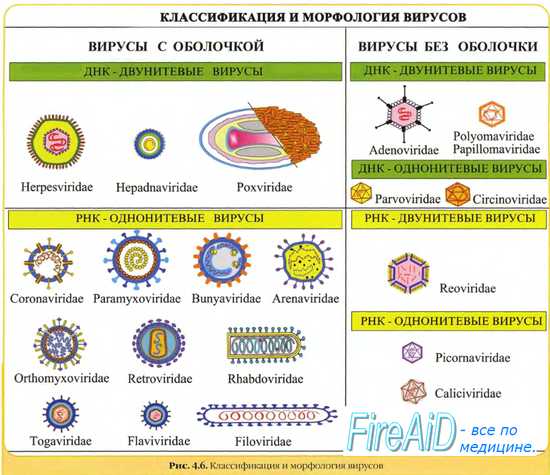
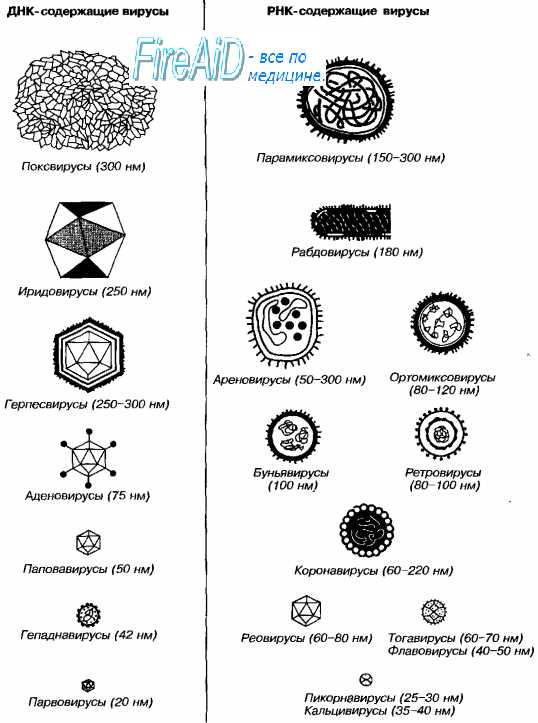
Морфология вирусов. Размеры вирусов.Несмотря на внутриклеточный паразитизм, среди вирусов имеются крупные виды, соизмеримые по размерам с микоплазмами и хламидиями. Например, вирус натуральной оспы достигает 400 нм и вполне сравним с риккетсиями (300-500 нм) и хламидиями (300-400 нм). По морфологии выделяют вирусы палочковидные (например, возбудитель лихорадки Эбола), пуле-видные (вирус бешенства), сферические (герпесвирусы), овальные (вирус оспы), а также бактериофаги, имеющие сложную форму (рис. 2-1). При всём разнообразии конфигураций, размеров и функциональных характеристик вирусам присущи некоторые общие признаки. В общем виде зрелая вирусная частица (вирион) состоит из нуклеиновой кислоты, белков и липидов, либо в его состав входят только нуклеиновые кислоты и белки. Нуклеиновые кислоты вирусовВирусы содержат только один тип нуклеиновой кислоты, ДИК или РНК, но не оба типа одновременно. Например, вирусы оспы, простого герпеса, Эпстайна-Барр — ДНК-содержащие, а тогавирусы, пикорнавирусы — РНК-содержащие. Геном вирусной частицы гаплоидный. Наиболее простой вирусный геном кодирует 3-4 белка, наиболее сложный — более 50 полипептидов. Нуклеиновые кислоты представлены однонитевыми молекулами РНК (исключая реовиру-сы, у которых геном образован двумя нитями РНК) или двухнитевыми молекулами ДНК (исключая парвовирусы, у которых геном образован одной нитью ДНК). У вируса гепатита В нити двухнитевой молекулы ДНК неодинаковы по длине. Вирусные ДНК образуют циркулярные, ковалентно-сцёпленные суперспирализованные (например, у паповавирусов) или линейные двухнитевые структуры (например, у герпес- и аденовирусов). Их молекулярная масса в 10-100 раз меньше массы бактериальных ДНК. Транскрипция вирусной ДНК (синтез мРНК) осуществляется в ядре заражённой вирусом клетки. В вирусной ДНК на концах молекулы имеются прямые или инвертированные (развёрнутые на 180″) повторяющиеся нуклеотидные последовательности. Их наличие обеспечивает способность молекулы ДНК замыкаться в кольцо. Эти последовательности, присутствующие в одно- и двух-нитевых молекулах ДНК, — своеобразные маркёры вирусной ДНК.
Вирусные РНК представлены одно- или двухнитевыми молекулами. Однонитевые молекулы могут быть сегментированными — от 2 сегментов у ареновирусов до 11 — у ротавирусов. Наличие сегментов ведёт к увеличению кодирующей ёмкости генома. Вирусные РНК подразделяют на следующие группы: плюс-нити РНК (+РНК), минус-нити РНК (-РНК). У различных вирусов геном могут образовывать нити +РНК либо -РНК, а также двойные нити, одна из которых -РНК, другая (комплементарная ей) — +РНК. Плюс-нити РНК представлены одиночными цепочками, имеющими характерные окончания («шапочки») для распознавания рибосом. К этой группе относят РНК, способные непосредственно транслировать генетическую информацию на рибосомах заражённой вирусом клетки, то есть выполнять функции мРНК. Плюс-нити выполняют следующие функции: служат мРНК для синтеза структурных белков, матрицей для репликации РНК, упаковываются в капсид с образованием дочерней популяции. Минус-нити РНК не способны транслировать генетическую информацию непосредственно на рибосомах, то есть они не могут функционировать как мРНК. Однако такие РНК служат матрицей для синтеза мРНК. Инфекционность нуклеиновых кислот вирусовМногие вирусные нуклеиновые кислоты инфекционны сами по себе, так как содержат всю генетическую информацию, необходимую для синтеза новых вирусных частиц. Эта информация реализуется после проникновения вириона в чувствительную клетку. Инфекционные свойства проявляют нуклеиновые кислоты большинства +РНК- и ДНК-содержащих вирусов. Двухнитевые РНК и большинство -РНК не проявляют инфекционных свойств. — Также рекомендуем «Капсид вируса. Функции капсида вирусов. Капсомеры. Нуклеокапсид вирусов. Спиральная симметрия нуклеокапсида. Кубическая симметрия капсида.» |
Источник
ирусы оспы. Виды вирусов оспы. Строение вирусов оспы.
Вирусы оспы. Виды вирусов оспы. Строение вирусов оспы.
Вирусы оспы (ВО) — самые крупные вирусы, содержащие ДНК, молекулярная масса которой больше, чем у любого другого вируса животных. Они широко распространены в природе, вызывают заболевания позвоночных и беспозвоночных с летальным исходом или в виде легко протекающей продолжительной инфекции с образованием доброкачественных опухолей. ВО передаются членистоногими при прямом контакте, аэрозольно или механически. ВО обычно имеют узкий круг хозяев, хотя некоторые из них представляют исключения, в том числе вирусы вакцины и оспы птиц. Вирусы оспы позвоночных включают восемь родов (орто-, пара-, ави-, капри-, лепори-, суи-, моллюсци- и ятапоксвирусы).
Разделение вирусов оспы позвоночных на роды проведено с учетом массы и структуры генома, размера и формы вириона, способности к генетической рекомбинации, антигенной связи и спектра патогенности.
Ортопоксвирусы. Типичный представитель — вирус осповакцины (ВОВ). Спектр естественных хозяев узкий, обычно ограничен одним видом животных: вирусы агглютинируют эритроциты цыплят; варьируют по вирулентности для естественных хозяев и лабораторных животных. Другие виды: вирусы оспы буйволов, верблюдов, крупного рогатого скота, мышей, обезьян и вирус натуральной оспы.
Парапоксвирусы. Типичный представитель — вирус контагиозного пустулёзного дерматита (эктимы) овец и коз (вирус орф). Представители рода серологически связаны между собой, но отличаются от представителей других родов; гемагглютинин не образуют. Другие виды: вирусы пустулёзного стоматита крупного рогатого скота, контагиозной эктимы серн и сыпи доярок.
Авипоксвирусы. Естественные хозяева — птицы; типичный представитель — вирус оспы кур. Члены рода серологически связаны между собой. Другие виды: вирусы оспы канареек, голубей, перепелов, воробьев, скворцов, индеек и оспы Юнко. Передаются членистоногими.
Каприпоксвирусы. Естественные хозяева — парнокопытные; типичный представитель — ВО мелких жвачных (овец и коз). В данный род входит также вирус кожной бугорчатки крупного рогатого скота.
Лепорипоксвирусы. Типичный представитель — вирус миксомы кроликов. Передается механически членистоногими. Другие виды: вирусы фибромы зайцев, кроликов (вирус Шоупа) и белок. Вирус злокачественной фибромы кроликов — летальный туморогенный поксвирус, возникший, по-видимому, вследствие рекомбинации между вирусами фибромы и миксомы кроликов.
Суипоксвирусы. Типичный представитель — вирус оспы свиней. В инфицированных клетках образуются несколько типов цитоплазматических включений и наблюдается вакуолизация ядра. Другие члены рода не определены.
Моллюсципоксвирусы. Типичный представитель — вирус контагиозного моллюска. Возможные члены рода — вирусы однокопытных и обезьян.
Ятапоксвирусы включают вирус опухолей обезьян Яба и вирус оспы Тана.
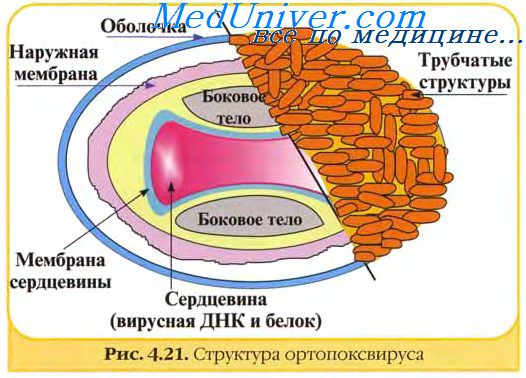
Вирусы оспы — наиболее крупные из всех вирусов животных. Под электронным микроскопом они выглядят как большие овальные (кирпичеобразные) частицы размером около 250-350×200-270 нм.
Орто-, ави-, лепорипоксвирусы более вытянуты, а вирус оспы свиней шире, чем другие ВО. Парапоксвирусы имеют овальную (коконообразную) форму и размер 260×160 нм.
В структуре вирусов оспы различают три основных компонента: двояковогнутую сердцевину, овальные боковые тела и оболочку вириона. Сердцевину вириона составляют ДНК и связанные с нею белки. Сердцевина окружена гладкой мембраной (толщиной около 5 нм), снаружи покрытой слоем вертикально уложенных и плотно прилегающих друг к другу цилиндрических субъединиц (5×10 нм). Вогнутость сердцевины с обеих сторон занята овальными образованиями (неизвестной природы), называемыми боковыми телами. Они как бы сдавливают сердцевину, придавая ей форму двояковогнутого диска, имеющего на разрезе вид гантели.
Вирионы заключены в липопротеиновую супероболочку, которая имеет толщину 20—30 нм и содержит липиды клетки и вирусспецифические белки.
Вирионы большинства вирусов оспы окружены слоем беспорядочно расположенных трубчатых структур, придающих им характерный вид. Эти структуры состоят из сферических субъединиц диаметром около 5 нм. Субъединицы построены из молекул протеина или гликопротеина. В состав ворсинок длиной 20 нм покрывающих поверхность вируса осповакцины (ВОВ), входит белок с молекулярной массой 58 кД, относящийся к главным полипептидам вириона. Поверхность парапоксвирусов покрыта длинными нитеподобными трубочками, уложенными крест-накрест, напоминающими клубок пряжи.
Вирус осповакцины содержит белки, липиды и ДНК, которые соответственно составляют 90, 5 и 3,2% массы вириона (5х10~15 г). В вирусе оспы птиц около 1/3 массы составляют липиды.
Вирион образуется включением ДНК внутрь незрелых вирусоподобных частиц, которые затем созревают, покрываясь дополнительно наружными оболочками. Репликация и сборка вирионов происходят в разных местах цитоплазмы (в виропластах или вирусных фабриках), и вирионы освобождаются почкованием или при лизисе клеток.
Вирионы, освободившиеся из клетки почкованием до ее разрушения, покрыты оболочкой, которая содержит клеточные липиды и несколько вирусспецифических белков.
— Также рекомендуем «Геном вирусов оспы. Антигены вирусов оспы.»
Оглавление темы «Частная вирусология.»:
1. ДНК-вакцины. Свойства и методы получения днк-вакцин.
2. Синтетические пептидные вакцины. Свойства синтетических пептидных вакцин.
3. Вирусы оспы. Виды вирусов оспы. Строение вирусов оспы.
4. Геном вирусов оспы. Антигены вирусов оспы.
5. Оспа овец. Оспа птиц. Особенности оспы овец и птиц.
6. Контагиозный пустулёзный дерматит. Эктима. Миксоматоз.
7. Асфаровирусы. Признаки и свойства асфаровирусов.
8. Иридовирусы. Герпесвирусы. Семейство герпесвирусов.
9. Эпидемиология герпесвирусов. Строение герпесвирусов.
10. Антигены вируса герпеса. Протеины герпесвирусов.
Источник
Вирусы – неклеточная форма жизни • Биология, Молекулярная биология | Фоксфорд Учебник
Вирус (от лат. virus — яд) — простейшая форма жизни, микроскопическая частица, представляющая собой молекулы нуклеиновых кислот (ДНК или РНК), заключенные в белковую оболочку (капсид) и способные инфицировать живые организмы.
Вирусы, за редким исключением, содержат только один тип нуклеиновой кислоты: либо ДНК, либо РНК (некоторые, например мимивирусы, имеют оба типа молекул).
Вирусы являются облигатными паразитами, так как не способны размножаться вне клетки. Вне клетки вирусные частицы ведут себя как химические вещества.
В настоящее время известны вирусы, размножающиеся в клетках растений, животных, грибов и бактерий (последних обычно называют бактериофагами). Обнаружены также вирусы, поражающие другие вирусы (вирусы-сателлиты).

Рис. 1 Бактериофаг
Впервые существование вируса (как нового типа возбудителя болезней) доказал в 1892 г. русский ученый Д. И. Ивановский. После многолетних исследований заболеваний растений табака в работе, датированной 1892 г., Д. И. Ивановский приходит к выводу, что табачная мозаика вызывается «бактериями, проходящими через фильтр Шамберлана, которые, однако, не способны расти на искусственных субстратах».
Пять лет спустя, при изучении заболеваний крупного рогатого скота, а именно ящура, был выделен аналогичный фильтрующийся микроорганизм. А в 1898 г., при воспроизведении опытов Д. Ивановского голландским ботаником М. Бейеринком, он назвал такие микроорганизмы «фильтрующимися вирусами». В сокращенном виде это название и стало обозначать данную группу микроорганизмов.
В 1901 г. было обнаружено первое вирусное заболевание человека — желтая лихорадка. Это открытие было сделано американским военным хирургом У. Ридом и его коллегами.
В 1911 г. Фрэнсис Раус доказал вирусную природу рака — саркомы Рауса (лишь в 1966 г., спустя 55 лет, ему была вручена за это открытие Нобелевская премия по физиологии и медицине).
В последующие годы изучение вирусов сыграло важнейшую роль в развитии эпидемиологии, иммунологии, молекулярной генетики и других разделов биологии. Так, эксперимент Херши и Чейза стал решающим доказательством роли ДНК в передаче наследственных свойств. В разные годы еще как минимум шесть Нобелевских премий по физиологии и медицине и три Нобелевских премии по химии были вручены за исследования, непосредственно связанные с изучением вирусов.
В 2002 г. в Нью-Йоркском университете был создан первый синтетический вирус (вирус полиомиелита).
Строение вирусов
Просто организованные вирусы состоят из нуклеиновой кислоты и нескольких белков, образующих вокруг нее оболочку — капсид. Примеров таких вирусов является вирус табачной мозаики. Его капсид содержит один вид белка с небольшой молекулярной массой.

Рис. 2 Вирус табачной мозаики
Сложно организованные вирусы имеют дополнительную оболочку — белковую или липопротеиновую; иногда в наружных оболочках сложных вирусов помимо белков содержатся углеводы. Примером сложно организованных вирусов служат возбудители гриппа и герпеса. Их наружная оболочка — это фрагмент ядерной или цитоплазматической мембраны клетки-хозяина, из которой вирус выходит во внеклеточную среду.

Рис. 3 Вирус гриппа
Распространение вирусов на Земле
Вирусы являются одной из самых распространенных форм существования органической материи на планете по численности: воды мирового океана содержат колоссальное количество бактериофагов (около 250 миллионов частиц на миллилитр воды), их общая численность в океане — около 4×, а численность вирусов (бактериофагов) в донных отложениях океана практически не зависит от глубины и всюду очень высока. В океане обитают сотни тысяч видов (штаммов) вирусов, подавляющее большинство которых не описаны и тем более не изучены. Вирусы играют важную роль в регуляции численности популяций некоторых видов живых организмов (например, вирус дикования раз в несколько лет сокращает численность песцов в несколько раз).
Генетические связи и происхождение вирусов
Вирусы имеют генетические связи с представителями флоры и фауны Земли. Согласно последним исследованиям, геном человека более чем на 32 % состоит из информации, кодируемой вирусподобными элементами и транспозонами. С помощью вирусов может происходить так называемый горизонтальный перенос генов (ксенология), то есть передача генетической информации не от непосредственных родителей к своему потомству, а между двумя неродственными (или даже относящимися к разным видам) особями. Так, в геноме высших приматов существует белок синцитин, который, как считается, был привнесен ретровирусом. Иногда вирусы образуют с животными симбиоз. Так, например, яд некоторых паразитических ос содержит структуры, называемые поли-ДНК-вирусами (Polydnavirus, PDV), имеющие вирусное происхождение.
Вирусы — сборная группа, не имеющая общего предка. В настоящее время существует несколько гипотез, объясняющих происхождение вирусов.
Считается, что крупные ДНК-содержащие вирусы происходят от более сложных (и, возможно, клеточных, таких как современные микоплазмы и риккетсии), внутриклеточных паразитов, утративших значительную часть своего генома. И действительно, некоторые крупные ДНК-содержащие вирусы (мимивирус, вирус оспы) кодируют функционально избыточные, на первый взгляд, ферменты, по-видимому, оставшиеся им в наследство от более сложных форм существования. Следует также отметить, что некоторые вирусные белки не обнаруживают никакой гомологии с белками бактерий, архей и эукариот, что свидетельствует о сравнительно давнем обособлении этой группы.
ДНК-содержащие бактериофаги и некоторые ДНК-содержащие вирусы эукариот, возможно, происходят от мобильных элементов — участков ДНК, способных к самостоятельной репликации в клетке.
Происхождение некоторых РНК-содержащих вирусов связывают с вироидами. Вироиды представляют собой высокоструктурированные кольцевые фрагменты РНК, реплицируемые клеточной РНК-полимеразой. Считается, что вироиды представляют собой «сбежавшие интроны» — вырезанные в ходе сплайсинга незначащие участки мРНК, которые случайно приобрели способность к репликации. Белков вироиды не кодируют. Считается, что приобретение вироидами кодирующих участков (открытой рамки считывания) и привело к появлению первых РНК-содержащих вирусов. И действительно, известны примеры вирусов, содержащих выраженные вироидподобные участки (вирус гепатита Дельта).
Вирусные частицы (вирионы) представляют собой белковую капсулу — капсид, содержащую геном вируса, представленный одной или несколькими молекулами ДНК или РНК. Капсид построен из капсомеров — белковых комплексов, состоящих, в свою очередь, из протомеров. Нуклеиновая кислота в комплексе с белками обозначается термином нуклеокапсид. Некоторые вирусы имеют также внешнюю липидную оболочку. Размеры различных вирусов колеблются от 20 (парвовирусы) до 500 (мимивирусы) и более нанометров. Вирионы часто имеют правильную геометрическую форму (икосаэдр, цилиндр). Такая структура капсида предусматривает идентичность связей между составляющими ее белками, и, следовательно, может быть построена из стандартных белков одного или нескольких видов, что позволяет вирусу экономить место в геноме.
Процесс вирусного инфицирования
Условно процесс вирусного инфицирования в масштабах одной клетки можно разбить на несколько взаимоперекрывающихся этапов:
• проникновение в клетку
• перепрограммирование клетки
• персистенция (переход в неактивное состояние)
• создание новых вирусных компонентов
• созревание новых вирусных частиц и их выход из клетки
ПРОНИКНОВЕНИЕ В КЛЕТКУ
На этом этапе вирусу необходимо доставить внутрь клетки свою генетическую информацию. Некоторые вирусы переносят также собственные белки, необходимые для ее реализации. Различные вирусы для проникновения в клетку используют разные стратегии: например, пикорнавирусы впрыскивают свою РНК через плазматическую мембрану, а вирионы ортомиксовирусов захватываются клеткой в ходе эндоцитоза, попадают в кислую среду лизосом, где происходит их окончательное созревание (депротеинизация вирусной частицы), после чего РНК в комплексе с вирусными белками преодолевает лизосомальную мембрану и попадает в цитоплазму. Вирусы также различаются по локализации их репликации, часть вирусов (например, те же пикорнавирусы) размножается в цитоплазме клетки, а часть (например, ортомиксовирусы) — в ее ядре.
ПЕРЕПРОГРАММИРОВАНИЕ КЛЕТКИ
При заражении вирусом в клетке активируются специальные механизмы противовирусной защиты. Зараженные клетки начинают синтезировать сигнальные молекулы — интерфероны, переводящие окружающие здоровые клетки в противовирусное состояние и активирующие системы иммунитета. Повреждения, вызываемые размножением вируса в клетке, могут быть обнаружены системами внутреннего клеточного контроля, и такая клетка должна будет «покончить жизнь самоубийством» в ходе процесса, называемого апоптозом или программируемой клеточной смерти. От способности вируса преодолевать системы противовирусной защиты напрямую зависит его выживание. Неудивительно, что многие вирусы (например, пикорнавирусы, флавивирусы) в ходе эволюции приобрели способность подавлять синтез интерферонов, апоптозную программу и т.д.
Кроме подавления противовирусной защиты вирусы стремятся создать в клетке максимально благоприятные условия для развития своего потомства.
ПРИМЕР ПЕРЕПРОГРАММИРОВАНИЯ СИСТЕМ КЛЕТКИ-ХОЗЯИНА
Хрестоматийным примером перепрограммирования систем клетки-хозяина является трансляция РНК энтеровирусов (семейство пикорнавирусы). Вирусная протеаза расщепляет клеточный белок eIF4G, необходимый для инициации трансляции подавляющего большинства клеточных мРНК (транслирующихся по так называемому кэп-зависимому механизму). При этом инициация трансляции РНК самого вируса происходит другим способом (IRES-зависимый механизм), для которого вполне достаточно отрезанного фрагмента eIF4G. Таким образом, вирусные РНК приобретают эксклюзивные «права» и не конкурируют за рибосомы с клеточными.
ПЕРСИСТЕНЦИЯ
Некоторые вирусы могут переходить в латентное состояние (так называемая персистенция для вирусов эукариот или лизогения для бактериофагов — вирусов бактерий), слабо вмешиваясь в процессы, происходящие в клетке, и активироваться лишь при определенных условиях. Так построена, например, стратегия размножения некоторых бактериофагов — до тех пор, пока зараженная клетка находится в благоприятной среде, фаг не убивает ее, наследуется дочерними клетками и нередко интегрируется в клеточный геном. Однако при попадании зараженной лизогенным фагом бактерии в неблагоприятную среду возбудитель захватывает контроль над клеточными процессами, так что клетка начинает производить материалы, из которых строятся новые фаги (так называемая литическая стадия). Клетка превращается в фабрику, способную производить многие тысячи фагов. Зрелые частицы, выходя из клетки, разрывают клеточную мембрану, тем самым убивая клетку. С персистенцией вирусов (например, паповавирусов) связаны некоторые онкологические заболевания.
СОЗДАНИЕ НОВЫХ ВИРУСНЫХ КОМПОНЕНТОВ
Размножение вирусов в самом общем случае предусматривает три процесса:
Транскрипция вирусного генома, то есть синтез вирусной мРНК.
Ее трансляция, то есть синтез вирусных белков.
Репликация вирусного генома (в некоторых случаях, когда генетическая информация вируса закодирована в виде РНК, геномная РНК одновременно играет роль мРНК, и, следовательно, процесс транскрипции в паразитируемой клетке не происходит за ненадобностью).
У многих вирусов существуют системы контроля, обеспечивающие оптимальное расходование биоматериалов клетки-хозяина. Например, когда вирусной мРНК накоплено достаточно, транскрипция вирусного генома подавляется, а репликация, напротив, активируется.
СОЗРЕВАНИЕ ВИРИОНОВ И ВЫХОД ИЗ КЛЕТКИ
В конце концов новосинтезированные геномные РНК или ДНК одеваются соответствующими белками и выходят из клетки. Следует сказать, что активно размножающийся вирус не всегда убивает клетку-хозяина. В некоторых случаях (например, ортомиксовирусы) дочерние вирусы отпочковываются от плазматической мембраны, не вызывая ее разрыва. Таким образом, клетка может продолжать жить и продуцировать вирус.
Источник