Проявляет свойства живых систем только в чужом организме вирус оспы
Содержание статьи
ВЕТРЯНАЯ ОСПА
Ветряная оспа — эта острая вирусная инфекция, характеризующаяся лихорадкой, толчкообразным появлением на коже и слизистой своеобразной пятнисто-везикулезной сыпи.
Возбудителем ветряной оспы является вирус крупных размеров (от 150 до 200 нм) из семейства Herpetosviridae. Этот вирус в большом количестве содержится в ветряночных пузырьках в первые 3-4 дня болезни, затем его количество быстро снижается, и после 7-го дня обнаружить его не удается.
В настоящее время установлена идентичность вируса, вызывающего опоясывающий герпес, и вируса ветряной оспы. Вирус имеет центральное ядро, липидную оболочку, содержит ДНК. Живет и размножается только в человеческом организме. К основным свойствам вируса относятся его летучесть и чрезвычайно малая устойчивость. Во внешней среде вирус быстро погибает, в капельках слизи, слюны сохраняется не более 10-15 мин; нагревание, солнечные лучи, УФ-излучение быстро инактивируют его.
Ветряная оспа встречается повсеместно и является чрезвычайно контагиозным заболеванием со 100-процентной восприимчивостью. Источник инфекции — больные ветряной оспой, иногда опоясывающим лишаем. Больные ветряной оспой становятся заразными в конце инкубационного периода (за 48 ч до появления сыпи) и продолжают представлять опасность для окружающих до 5-го дня с момента появления последнего элемента сыпи. Механизм передачи — воздушно-капельный, при этом вирус выделяется в огромном количестве при кашле, разговоре, чихании. Возможен вертикальный механизм передачи вируса плоду от матери, болевшей ветряной оспой в период беременности. Ветряной оспой болеют преимущественно дети в возрасте от 1 до 10 лет, причем пик заболеваемости приходится на 3-4 года.
Входными воротами инфекции является слизистая оболочка верхних дыхательных путей. Размножаясь на слизистых оболочках, вирус в конце инкубационного периода попадает в кровь, обусловливая вирусемию. С током крови он разносится по всему организму. Возбудитель фиксируется преимущественно в эпителии кожи и слизистых оболочек, где, размножаясь, приводит к появлению патогномоничной для ветряной оспы сыпи.
Вирус не только дерматотропен, но,в известной степени, и нейротропен, что проявляется в его способности вызывать поражения со стороны нервной системы. Пузырьки при ветряной оспе образуются в результате вакуолизации клеток с очень незначительной сетчатостью. На ранних стадиях ядра пораженных клеток содержат сферические эозинофильные включения (тельца Тиццера). В процессе дегенерации ядер эти тельца выходят в цитоплазму. Клеточные вакуоли быстро сливаются со смежными, образуя везикулу. Вначале везикула бывает многокамерной, состоит из маленьких полостей, отделяемых друг от друга клеточными тяжами, которые в дальнейшем быстро разрываются благодаря наполнению везикул жидкостью. Поражается в основном мальпигиевый слой. Изменения на слизистых оболочках носят тот же характер, что и в эпидермисе. Элементы сыпи на слизистых оболочках при ветряной оспе не оставляют после себя рубцов, так как некроз эпителия при ветряной оспе обычно не проникает глубже герминативного слоя.
В течении ветряной оспы выделяют 4 периода: инкубационный, продромальный, периоды высыпания и образования корочек. Инкубационный период составляет при ветряной оспе 10-21 день. Продромальные явления могут отмечаться в течение 1-2 сут до начала высыпания. При этом больной испытывает недомогание, снижается аппетит, возникают головная боль, тошнота, иногда рвота. Если продромальный период отсутствует, то заболевание начинается с появления сыпи. Период высыпания у большинства больных протекает без особых нарушений общего состояния. Лихорадка совпадает с периодом массового появления сыпи, при этом у взрослых она достигает значительных цифр. Высыпания появляются толчкообразно, поэтому лихорадка может носить волнообразный характер.
Первые элементы сыпи могут появиться на любом участке тела, но чаще на лице, волосистой части головы, спине, реже — на животе, груди, плечах, бедрах. На ладонях и подошвах сыпь, как правило, отсутствует.
Клинический пример: больной С.,23 года, студент, обратился к участковому врачу с жалобами на повышение температуры до 38,8°С , головную боль, слабость, недомогание, высыпания на лице, волосистой части головы, туловище и конечностях. Со слов пациента, он заболел два дня назад; возникли недомогание, слабость, температура повысилась до 37,4°С. На следующий день состояние ухудшилось, температура поднялась до 38,0°С, отмечалась головная боль, была однократно рвота. К вечеру появилась обильная сыпь (рис. 1).
 |
| Рисунок 1. Полиморфная сыпь при ветряной оспе на лице и волосистой части головы |
Объективно: состояние средней тяжести. Температура тела — 39,0°С. На коже лица, волосистой части головы, туловище, конечностях имеется обильная полиморфная сыпь. На одном участке можно встретить папулы, везикулы, пустулы. Беспокоит кожный зуд. Слизистая ротоглотки гиперемирована, на мягком небе и язычке отмечаются в небольшом количестве везикулы и эрозии. Пальпируются увеличенные болезненные лимфатические узлы шеи. В легких дыхание везикулярное, хрипов нет. ЧДД — 22/мин. Тоны сердца приглушены, ритм правильный. ЧСС — 92/мин. АД — 90/60 мм рт. ст. Язык сухой, обложен белым налетом. Живот при пальпации мягкий, безболезненный во всех отделах. Печень, селезенка не увеличены. Мочевыделение не нарушено. Менингеальной, очаговой неврологической симптоматики нет. В группе, где учится пациент, отмечались подобные случаи болезни.
Учитывая острое начало болезни, наличие лихорадки, симптомов интоксикации, а также полиморфной сыпи на волосистой части головы, лице, туловище, конечностях и слизистой ротоглотки(рис. 2), пациенту можно поставить предварительный диагноз «ветряная оспа».
|
| Рисунок 2. Эрозия на слизистой ротоглотки |
Элементы ветряной оспы проходят следующие стадии развития: пятна, папулы, везикулы, корочки. Вначале появляется красное пятнышко размером от булавочной головки до чечевичного зерна круглой или овальной формы. В течение нескольких часов пятна приобретают характер папул с четко очерченным контуром. Через несколько часов либо на следующий день в центре элементов образуется пузырек-везикула с ровными краями и прозрачным серозным содержимым. В случае нагноения везикул образуются пустулы, при этом отмечается глубокое поражение кожи, что может приводить к образованию небольших западений в центре пустул, которые оставляют рубцы. Нередко отмечаются высыпания на конъюнктивах глаз, слизистой оболочке рта (твердом небе, слизистой оболочке щек, деснах, язычке, задней стенке глотки), иногда гортани и половых органах. Ветряночные пузырьки напоминают афты, однако везикулы быстро повреждаются, образуя небольшие поверхностные язвочки (эрозии). Для ветряной оспы характерны многократные высыпания, появляющиеся в несколько приемов, последовательно, в течение 2-5 дней. Такое волнообразное высыпание приводит к тому, что на одном и том же ограниченном участке кожи отмечаются элементы ветряной оспы, находящиеся на различных этапах развития, что придает ветряной оспе полиморфный характер. Помимо специфического высыпания в некоторых случаях в продромальном периоде ветряной оспы отмечается появление на коже продромальной сыпи, английское ее название «rash» («реш»), которая предшествует ветряночному высыпанию. Продромальная сыпь обычно появляется на груди, изредка на верхних конечностях, реже — на лице. Она не обильна, эфимерна, бывает скарлатиноподобной, кореподобной, реже носит геморрагический или эритематозный характер.
В зависимости от течения различают следующие клинические формы ветряной оспы.
Типичные и атипичные. Среди атипичных, в свою очередь, различаются следующие формы: рудиментарная, геморрагическая, буллезная, гангренозная, генерализованная.
По тяжести формы заболевания могут быть легкие, средние и тяжелые. Последние бывают: а) с выраженной общей интоксикацией; б) с выраженными изменениями на коже.
При типичной форме общее состояние больного страдает редко, температура кратковременно повышается до 37-38°С, обычно в период массового высыпания. Полиморфизм сыпи хорошо выражен. Общая продолжительность высыпания при типичной форме составляет 3-5 дней. Энантема наблюдается в 70% случаев. Осложнения встречаются редко.
При атипичном течении ветряной оспы отклонения от обычного возможны как в стороны легких (рудиментарных), так и редко встречающихся тяжелых форм болезни с необычными кожными проявлениями, нередко с летальным исходом.
Рудиментарная форма протекает без сыпи или со скудным высыпанием, при этом элементы сыпи не достигают своего полного развития, ограничиваясь только появлением мелких красных пятнышек. Иногда сыпь может состоять всего лишь из нескольких папул и мелких, едва заметных пузырьков, при этом полиморфизм сыпи слабо выражен. Высыпания на слизистых также единичны.
При геморрагической форме на 2-3- день болезни высыпания наблюдается скопление геморрагического содержимого в пузырьках при одновременном появлении петехий и крупных кровоизлияний на коже и слизистых оболочках. Возможны кровотечения из десен, носа и желудочно-кишечного тракта. Корочка, образующаяся при этой форме, черного цвета, глубоко находящаяся в коже, нередко изъязвляется (рис. 3).
|
| Рисунок 3. Полиморфные высыпания с геморрагическим компонентом у больного ветряной оспой |
Эта форма болезни наблюдается у больных с предшествующими геморрагическими явлениями — при капилляротоксикозе, болезни Верльгофа.
При буллезной форме на коже наряду с типичными ветряночными пузырьками наблюдаются большие дряблые, тонкостенные пузырьки с желтовато-мутным содержимым, которые подсыхают медленнее, чем обычно, и образуют корочку или же, лопаясь, превращаются в длительно не заживающие мокнущие поверхности. При буллезной форме самостоятельно образованные пузырьки, быстро увеличиваясь по периферии, могут сливаться в большие пузыри.
Для гангренозной формы характерен прогрессирующий некроз. Через несколько дней после появления везикул вокруг некоторых из них появляются гангренозные ободки, распространяющиеся по периферии. Пузырьки круглые, крупные, диаметром в несколько сантиметров, наполненные гнойно-кровянистым содержимым. После вскрытия на их месте образуется некротический струп, при отторжении которого обнаруживаются долго не заживающие язвы различной глубины с подрытыми краями, с грязным гнойным дном. Для этой формы характерна выраженная интоксикация, высокая летальность. Гангренозная форма наблюдается редко, главным образом у детей со сниженной реактивностью организма.
Генерализованная (висцеральная) форма возникает при лечении стероидными гормонами, у людей, ослабленных тяжелыми заболеваниями. Характерно поражение внутренних органов. Течение болезни очень тяжелое, нередко с летальным исходом.
Осложнения
Ветряная оспа считается доброкачественным заболеванием, однако в ряде случаев могут развиться осложнения (примерно у 5% больных), иногда очень тяжелые.
Среди осложнений ветряной оспы наиболее часты различные гнойные поражения кожи — абсцессы, фурункулы, флегмоны. В основе гнойных осложнений кожи лежит суперинфекция стафилококками или стрептококками. Одной из частых причин инфицирования является расчесывание больным зудящих мест кожи. В большинстве случаев входными воротами гнойной инфекции становится поврежденный ветряночный пузырек.
Менее распространенные осложнения: энцефалит, миокардит, пневмония, кератит, нефрит, артрит, гепатит.
Клинически выраженная картина ветряночной пневмонии — одна из особенностей ветряной оспы взрослых. При этом непосредственно перед высыпанием, чаще одновременно с массовым высыпанием и повышением температуры, у больного появляются одышка, цианоз, кашель с кровянистой мокротой, боли в груди. Физикальные изменения часто отсутствуют или очень незначительны. На рентгенограмме видны обильные мелкие очажки на всем протяжении. Клинические проявления пневмонии наблюдаются в течение 7-10 дней, рентгенологические изменения — до 1-2 месяцев.
Поздние ветряночные энцефалиты появляются чаще во время угасания сыпи с 5-15-го дня болезни, но могут возникать и позже. Для поздних ветряночных энцефалитов обычны более плавное развитие и независимость от характера высыпания и тяжести течения ветряной оспы. Одновременно с кратковременными подъемами температуры у больных к концу ветряночного высыпания наблюдаются вялость, головная боль, рвота, а через несколько дней основные симптомы. Энцефалиты могут быть различной локализации: атактические формы, миелиты и энцефаломиелиты, энцефалиты с поражением психики, изолированное поражение нервов, менингоэнцефалиты, при этом чаще регистрируется атактический синдром. Наблюдались поздние ветряночные энцефалиты с парезом конечностей и полной временной потерей зрения. Среди энцефалитов, дающих менее благоприятный прогноз, можно упомянуть формы с поражениями подкорковых узлов. К исключительно редким последствиям ветряночных менингоэнцефалитов относится поражение психики с последующей идиотией. Для ветряночного энцефалита и других неврологических осложнений ветряной оспы характерна обратимость процесса.
Поражение почек — редкое осложнение. Нефрит при ветряной оспе чаще появляется на 2-й неделе к концу высыпания. Он начинается остро: с подъема температуры до высоких показателей, рвоты, головной боли и болей в животе. Изменения в моче бывают умеренными и выражаются в появлении незначительного количества белка, небольшой гематурии, единичных цилиндров. Это осложнение протекает легко и к 12-15-му дню болезни исчезает.
Ветряная оспа представляет значительную опасность для будущих матерей, особенно в первые месяцы беременности. Возможно патологическое влияние на развитие плода, иногда выкидыш, в поздние сроки могут возникнуть тяжелые висцеральные формы ветряной оспы у новорожденных, способные привести к летальному исходу.
Диагностика ветряной оспы в типичных случаях не представляет затруднений. Диагноз устанавливается, главным образом, на основании клиники, при этом учитываются данные эпидемиологического анамнеза. Из лабораторных методов используют вирусоскопический, вирусологический, молекулярно-биологический и серологический. Вирусоскопический метод заключается в окрашивании содержимого пузырька серебрением, вирус обнаруживается с помощью обычного светового микроскопа.
Основным вирусологическим методом можно считать ПЦР, так как она очень информативна. Этим способом определяют ДНК-вируса. Кроме того, можно выделить вирус в культуре тканей, но данный метод очень дорогостоящий и занимает много времени. Из серологических методов используют РСК, РИМФ и ИФА.
Дифференциальная диагностика
Ветряную оспу следует дифференцировать с натуральной оспой, опоясывающим герпесом, полиморфной экссудативной эритемой, импетиго, везикулезным риккетсиозом (табл. 1).
Лечение
Большое место в терапии больного занимают гигиенические мероприятия, цель которых — предотвращение вторичной инфекции. В период высыпания и лихорадки требуется соблюдение постельного режима. Элементы сыпи смазывают раствором анилиновых красителей: 1-2% водным или спиртовым растворами бриллиантового зеленого, генцианового фиолетового, метиленового синего. Очень хорошо смазывать элементы сыпи водными растворами марганцовокислого калия в разведении 1:5000 или 2-3% йодной настойки. При поражении слизистых полости рта необходимо полоскание слабым раствором марганцовокислого калия. Афты смазывают раствором бриллиантового зеленого. Эффективна обработка 3% раствором перекиси водорода. Для уменьшения зуда кожу можно смазывать глицерином, обтирать водой с уксусом или спиртом. Показаны антигистаминные препараты (псило-бальзам, фенистил). При тяжелой форме заболевания с выраженными симптомами интоксикации проводится дезинтоксикационная терапия. Эффективны в отношении лечения ветряной оспы противовирусные препараты (ацикловир, видарабин) (табл. 2). Антибиотики при ветряной оспе назначают в случае присоединения вторичной бактериальной инфекции и гнойных осложнений.
Общая профилактика ветряной оспы заключается в своевременной диагностике, ранней изоляции больного с последующим проветриванием и влажной уборкой помещения. Изоляция больного прекращается через 5 дней после появления последнего свежего элемента сыпи. Дети до 7 лет, вступавшие в контакт с больными ветряной оспой, не болевшие и посещающие детские учреждения, изолируются до 21-го дня с момента контакта.
Литература
- Зубик Т. М. и др. Дифференциальная диагностика инфекционных болезней. — Л., 1991.
- Руководство по инфекционным болезням // Под ред. Ю. В. Лобзина. — СПб., 2001.
- Юнусова Х. А., Шамсиев Ф. С. Ветряная оспа. — М., 1999.
- Баран В. М., Талапин В. И. Фармакотерапия инфекционных болезней. — Минск, 1995.
- Детские инфекционные болезни // Под ред. В. В. Фомина. — Екатеринбург, 1992.
- Беренбейн Б. А. и др. Дифференциальная диагностика кожных болезней. — М., 1989.
- Соринсон С. Н. Неотложные состояния у инфекционных больных. — Л.: Медицина, 1990.
Т. К. Кускова, кандидат медицинских наук
Е. Г. Белова, кандидат медицинских наук
Т. Э. Мигманов, кандидат медицинских наук
Московский медико-стоматологический университет
Обратите внимание!
- К основным свойствам вируса относятся его летучесть и чрезвычайно малая устойчивость.
- Ветряная оспа встречается повсеместно и является чрезвычайно контагиозным заболеванием со 100-процентной восприимчивостью. Источник инфекции — больные ветряной оспой, иногда опоясывающим лишаем.
- Возможен вертикальный механизм передачи вируса плоду от матери, болевшей ветряной оспой в период беременности.
- Элементы сыпи на слизистых оболочках при ветряной оспе не оставляют после себя рубцов, так как некроз эпителия при ветряной оспе обычно не проникает глубже герминативного слоя.
- Инкубационный период при ветряной оспе составляет 10-21 день.
- Период высыпания у большинства больных протекает без особых нарушений общего состояния.
Источник
Вирусные геномы в системе эволюции
Статья на конкурс «био/мол/текст»: Изучение виромов (популяций вирусов, объединенных общей средой обитания) практикуется в современной вирусологии всё чаще. Подобный подход приносит неожиданные результаты — концентрация вирусов в природе, оказывается, чрезвычайно высока. Например, в морской среде количество вирусных частиц превосходит количество клеток в разы. При этом вирусные частицы способны к передаче генетической информации. Исследования, опубликованные в 2010 году, показали, что агенты переноса генов (АПГ, особая разновидность вирусоподобных частиц, участвующих в горизонтальном переносе генов) привносят в геном бактерий новые фрагменты значительно чаще, чем считалось ранее. А если вирусные частицы настолько распространены в биосфере, и большая часть из них способна к переносу генов, то важно рассмотреть вирусы в рамках общей модели эволюции.
Вопрос о происхождении вирусов
Существует три основные теории возникновения вирусов:
- Сценарий «сбежавших генов», приравнивающий вирусы к генам клеточных организмов, которые «выпали» из клеточного генома и переключились на «эгоистичный» режим, — теория клеточного происхождения.
- Дегенерация одноклеточных паразитов до вирусного состояния — регрессивная гипотеза.
- Происхождение вирусов из первичных генетических элементов — гипотеза доклеточного происхождения.
Однако фундаментальные достижения, такие как открытие рибозимов (молекул РНК, обладающих каталитическим действием), разработка гипотезы о «мире РНК» [2], открытие новых форм вирусов и успехи в геномике делают наименее популярную в прошлом гипотезу доклеточного происхождения все более весомой. И то, что некоторые вирусные белки не обнаруживают гомологии с белками бактерий, архей и эукариот [3], свидетельствует о сравнительно давнем обособлении этой группы. В остальном же достоверно объяснить происхождение вирусов на основании трёх закрепившихся классических гипотез пока еще не вполне удаётся.
Возникновение паразитов — неизбежное последствие эволюционного процесса
Если рассматривать эволюцию как единый процесс, становится понятно, что виды никогда не эволюционируют независимо друг от друга. Более того, паразиты появляются и в компьютерных моделях эволюции простых репликаторов (самовоспроизводящихся РНК-подобных систем) [9]. Так исследования, проведенные в 2008 году Набуто Такеучи и Паулин Хогвег, представляют собой расчет такой репликативной системы [12]. Простота системы позволяет явно смоделировать отображение генотип-фенотип взаимодействий отдельных репликаторов. Результаты показали, что популяция репликаторов, первоначально состоящая из одного генотипа, развивается в сложную экосистему до четырех «видов». В этой диверсификации «виды» эволюционируют путем приобретения уникальных генотипов с отличной экологической функциональностью, причем два из четырех «видов» становятся паразитами, неспособными к самостоятельной репликации.
Анализ диверсификации показывает, что паразитарные репликаторы, которые были придуманы, чтобы дестабилизировать разнообразие репликатора, на самом деле способствуют эволюции разнообразия посредством создания новой «ниши» для каталитических репликаторов (самовоспроизводящихся репликаторов). Это также делает существующую систему репликатора чрезвычайно стабильной при эволюции паразитов, поскольку в такой системе паразиты имеют преимущество репликации на катализаторах.
Дело в том, что репликация катализаторов требует некоторого периода времени, и паразиты, которые не копируют другие молекулы, могут тратить больше времени на репликацию, чем катализаторы. Таким образом, присутствие катализаторов влечет за собой «нишу» для паразитов. В нынешней системе (рис. 4), C-катализатор (cat-C) создает такую нишу, и это позволяет эволюционировать G-паразиту (par-G). Кроме того, как только создается структура C-катализатор-G-паразит, она создает еще один нишу для фенотипа, который может высвободиться из G-паразита. И если такой фенотип приобретает А-катализатор (cat-A), он тоже эволюционирует. Наконец, создание такого альтернативного катализатора создает нишу для фенотипа, который мог бы паразитировать на этом альтернативном катализаторе (par-U).
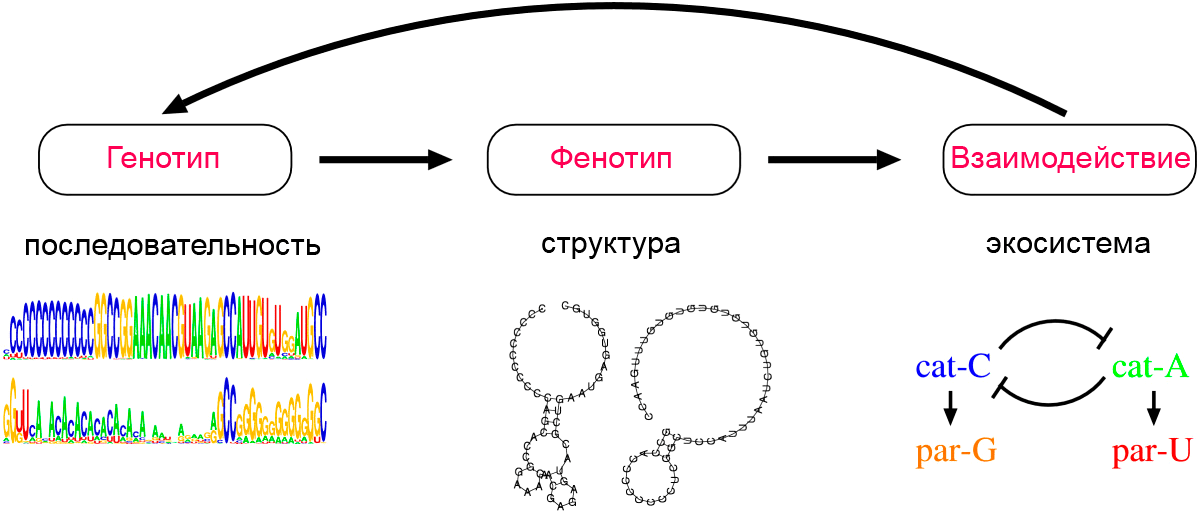
Рисунок 4. Схематическое представление структуры модели эволюции РНК-подобной системы. На втором этапе цепочки последовательностей начинают соединяться комплементарными связями сами с собой. В результате у двух видов (cat-C и cat-A) возникает вторичная структура молекулы, которая обладает каталитическим свойством. Она ускоряет собственную репликацию (или репликацию несвернувшихся соседей). Два вида при этом приобретают паразитические свойства (par-G и par-U). Пояснения в тексте.
Таким образом, паразитарные репликаторы способствуют эволюции разнообразия, вместо того, чтобы мешать этому разнообразию. Это также делает существующую систему репликатора чрезвычайно стабильной при эволюции паразитов.
Согласно гипотезе Черной Королевы, чтобы поддержать свое существование в постоянно эволюционирующем мире, вид должен реагировать на эти эволюционные изменения и должным образом приспосабливаться к среде. Поэтому, если мы говорим о вирусах как о паразитах, мы обязаны представлять себе взаимоотношения вируса с хозяином. В борьбе с вирусом хозяева развивают новые защитные механизмы, а паразиты отвечают, развивая механизмы для атаки и взлома защиты. Этот процесс может длиться бесконечно либо до вымирания одной из противоборствующих сторон. Так множественные системы защиты составляют существенную часть геномов всех клеточных организмов, а взлом защиты — одна из основных функций генов у вирусов с большими геномами .
Понятие о вирусном геноме, типы вирусных генов, концепция генов-сигнатур
В исследовании, проведенном вирусологом Евгением Куниным и его коллегами [16], анализ последовательностей вирусных геномов выявил несколько категорий вирусных генов, принципиально отличающихся по происхождению. Можно обсуждать, какая степень дробности классификации оптимальна, но четко различаются пять классов, укладывающихся в две более крупные категории.
Гены с четко опознаваемыми гомологами у клеточных форм жизни:
- Гены, присутствующие у узких групп вирусов (обычно это гены, гомологичные генам хозяев этих вирусов).
- Гены, консервативные среди большой группы вирусов или даже нескольких групп и имеющие относительно отдаленные клеточные гомологи.
Вирус-специфичные гены:
- «Бесхозные» (orphan) гены — гены без выявленных гомологов, кроме как у близкородственных вирусов.
- Вирус-специфические гены, консервативные для относительно широкой группы вирусов, но не имеющие гомологов у клеточных форм жизни.
- Гены — вирусные сигнатуры.
Вирусные сигнатуры — это гены, общие для многих разнообразных групп вирусов, имеющие лишь отдаленные гомологи у клеточных организмов, для которых имеются убедительные свидетельства в пользу монофилии (общего происхождения) всех вирусных членов соответствующих семейств генов [9]. Фактически эти гены являются признаками «вирусного состояния». Гены-сигнатуры предшествуют клеткам и происходят непосредственно из первичного, доклеточного пула генов.
Таким образом, отличительные особенности генов-сигнатур:
- Происхождение из первичного пула генов;
- Наличие лишь очень отдаленных гомологов среди генов клеточных форм жизни, из чего можно сделать вывод, что они никогда не входили в геномы клеточных форм;
- Необходимость для репродукции вирусов.
Из всего вышесказанного следует, что эти гены переходили от вируса к вирусу (или к элементу, подобному вирусу) на протяжении четырех миллиардов лет эволюции жизни, а вирусные геномы появились благодаря перемешиванию и подгонке друг к другу генов в гигантской генетической сети, которую представляет собой мир вирусов. Многочисленные гены клеточных форм жизни также пронизывают эту сеть, прежде всего благодаря геномам крупных вирусов, таких как NCDLV и крупным бактериофагам, которые позаимствовали множество генов от своих хозяев на разных этапах эволюции. Однако большинство заимствованных генов сами по себе не критичны для репликации и экспрессии вирусного генома (исключая некоторые случаи возможного неортологичного замещения генов-сигнатур); обычно эти гены участвуют во взаимодействии между вирусом и хозяином. Таким образом, несмотря на интенсивный взаимообмен генами с хозяевами, вирусы всегда происходят от других вирусов.
Вирусы, встроенные в геном, и горизонтальный перенос генов
В процессе эволюции многие вирусы встроились в геномы клеточных форм жизни путем горизонтального переноса генов (ГПГ). Впервые горизонтальный перенос был описан в 1959 году, когда ученые продемонстрировали передачу резистентности к антибиотикам между разными видами бактерий. В 1999 году Рави Джайн, Мария Ривера и Джеймс Лейк в своей статье писали о произошедшей значительной передаче генов между прокариотами [17]. Этот процесс, по-видимому, оказал некоторое влияние также и на одноклеточные эукариоты. В 2004 году Карл Вёзе опубликовал статью, в которой утверждал, что между древними группами живых организмов происходил массивный перенос генетической информации. В древнейшие времена преобладал процесс, который он называет горизонтальным переносом генов. Причем, чем дальше в прошлое, тем это преобладание сильнее [18].
Современные же данные показывают, что ГПГ имеет огромное влияние не только на эволюцию простейших, но и на эволюцию многоклеточных. Молекулярный биолог Петер Гогартен описал горизонтальную генную передачу как «новую парадигму биологии».
Горизонтальный перенос генов — процесс, в котором организм передаёт генетический материал другому организму, не являющемуся его потомком. Горизонтальная передача генов реализуется через различные каналы генетической коммуникации — процессы конъюгации, трансдукции, трансформации, переноса генов в составе плазмидных векторов, вирусов, мобильных генетических элементов (МГЭ).
Трансдукция — перенос бактериофагом (агентами переноса генов, АПГ) в заражаемую клетку фрагментов генетического материала клетки, исходно содержавшей бактериофаг [19]. Такой бактериофаг обычно переносит лишь небольшой фрагмент ДНК хозяина от одной клетки (донор) к другой (реципиент). В зависимости от типа трансдукции — неспецифической (общей), специфической или абортивной, геном фага или хозяина-бактерии может быть изменен тем или иным образом:
- При неспецифической трансдукции (рис. 5) ДНК клетки-хозяина включаются в частицу фага (дополнительно к его собственному геному или вместо него);
- При специфической трансдукции гены фага замещаются генами хозяина;
- При абортивной трансдукции внесённый фрагмент ДНК донора не встраивается в ДНК хозяина-реципиента, а остаётся в цитоплазме и не реплицируется. Это приводит к тому, что при клеточном делении он передаётся только одной из дочерних клеток и затем теряется в потомстве.
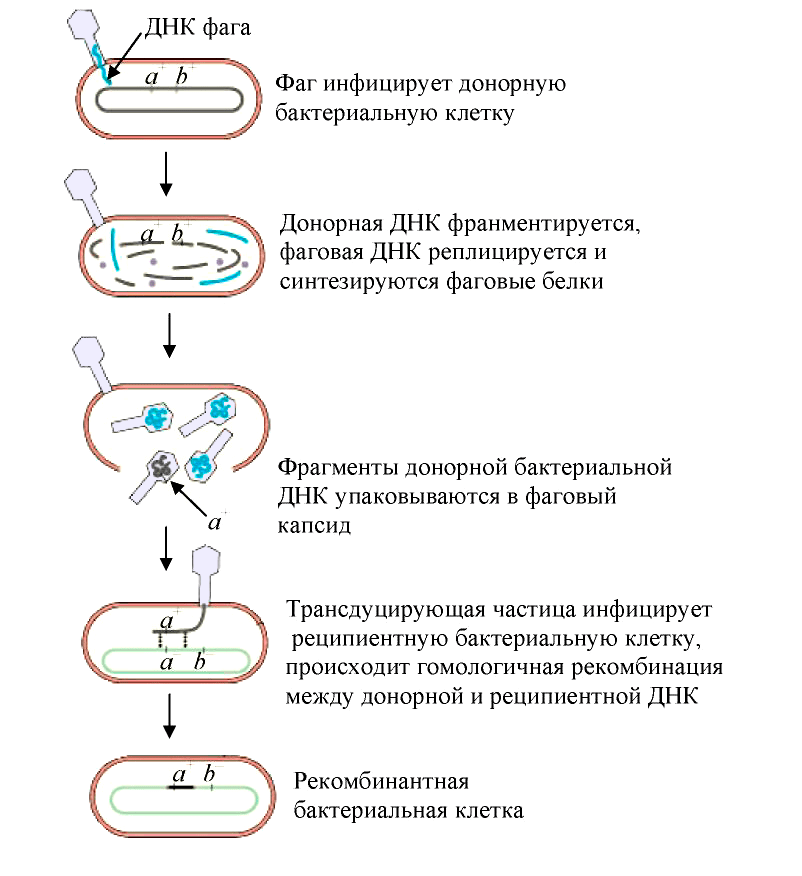
Рисунок 5. Схема общей трансдукции
Наиболее известным примером специфической трансдукции служит трансдукция, осуществляемая фагом λ. Поскольку этот фаг при переходе в состояние профага включается в хромосому бактерий между генами, кодирующими синтез галактозы и биотина, именно эти гены он может переносить при трансдукции.
Важнейшую роль в эволюции животных играют гены, которые животные заимствуют у вирусов, транспозонов, ретротранспозонов, дельта-ретровирусов (экозогенные и эндогенные вирусы). Фрагменты ДНК вирусов и транспозонов часто «приручаются» высшими организмами и начинают выполнять полезные функции в геноме. Большинство встроенных (ретровирусных, если говорить о высших позвоночных) геномов совершенно бесполезны для хозяина. Фактически идет постоянный процесс превращения так называемой «мусорной» или «эгоистической» ДНК вирусного происхождения в полезные элементы генома. Явление это настолько широко распространено, что для него даже предложен специальный термин — молекулярное одомашнивание [21].
Вот несколько примеров важных эволюционных событий, связанных с молекулярным одомашниванием:
- Ферменты теломеразы, служащие для восстановления концевых участков хромосом, возможно, ведут свое происхождение от обратных транскриптаз, кодируемых ретровирусами и ретротранспозонами [22];
- Белки RAG, играющие ключевую роль в системе адаптивного иммунитета, по-видимому, происходят от прирученных транспозаз — ферментов, кодируемых транспозонами;
- Ген Peg10, необходимый для развития плаценты, был позаимствован древними млекопитающими у ретротранспозона (рис. 6) [23].
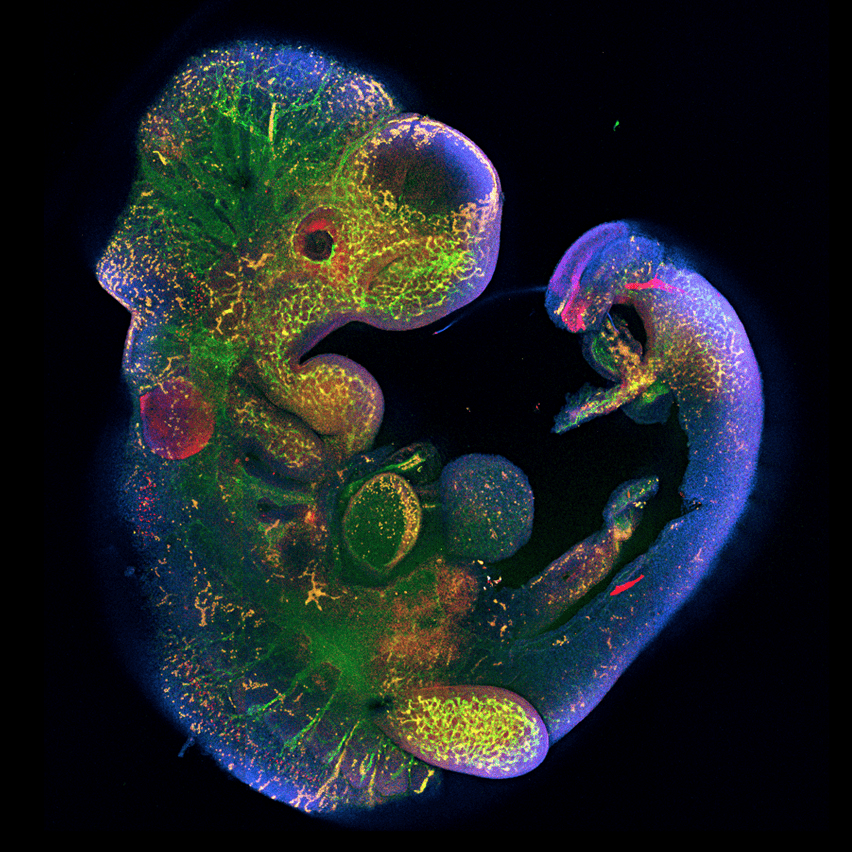
Рисунок 6. Роль гена Peg10 в эмбриональном развитии. Ученые под руководством Рюичи Оно из Токийского медицинского университета Японии показали, что у мышей с выключенным геном Peg10 нарушается развитие плаценты, от чего эмбрион погибает через 10 дней после зачатия [24].
В 2008 году в ходе целенаправленного поиска неиспорченных вирусных генов в геноме человека исследователи нашли два очень похожих друг на друга ретровирусных гена (их назвали ENVV1 и ENVV2), которые, по всей видимости, находятся в рабочем состоянии [25]. Это гены белков оболочки ретровируса. Каждый из них входит в состав своего эндогенного ретровируса (ЭРВ), причем все остальные части этих ЭРВ давно не функционируют.
Авторы нашли гены ENVV1 и ENVV2 и в геномах других приматов: у человекообразных (шимпанзе, орангутана) и других обезьян Старого Света (макаки-резуса, зеленой мартышки), а также у обезьян Нового Света — мармозетки и беличьей обезьяны. Однако у наших более отдаленных родственников (лемуры) этих генов нет. Сопоставив данные по нуклеотидным последовательностям генов ENVV1 и ENVV2 у разных обезьян, исследователи смогли реконструировать эволюционную историю этих генов. В эволюции приматов имело место приобретение полезных генов «со стороны», а именно от ретровирусов. По своему происхождению они являются генами белков оболочки ретровируса [21]. Исходный ретровирус встроился в геном наших предков и стал эндогенным ретровирусом после того, как разделились линии обезьян и лемуров, но до того, как разошлись обезьяны Старого и Нового света, то есть примерно 43-77 млн. лет назад. Этот ЭРВ вскоре подвергся двум последовательным дупликациям, и в результате получилось три одинаковых ЭРВ, расположенных по соседству на одной хромосоме. Все участки этих ЭРВ, кроме генов белков оболочки, стали постепенно дегенерировать. Ген ENVV2 приобрел в результате мутаций какое-то полезное для хозяина свойство и стал сохраняться отбором. Другие два гена, по-видимому, тоже приобрели полезные функции, но они оказались не столь незаменимыми, как ENVV2. Поэтому в некоторых эволюционных линиях обезьян эти гены были утрачены или выведены мутациями из строя. В частности, общий предок человека и шимпанзе потерял ген ENVV3.
Вирусные гены ENVV1 и ENVV2 у человека и обезьян работают в плаценте и, скорее всего, выполняют следующие функции:
- Управление слиянием клеток в ходе формирования наружного слоя плаценты (синцитиотрофобласт);
- Защита эмбриона от иммунной системы матери (у обоих белков есть участок, обладающий свойством искусственного угнетения иммунитета, это вполне объясняется тем, что изначально они входили в состав вирусной оболочки);
- Защита эмбриона от «диких» ретровирусов. У ENVV1 и ENVV2 сохранились участки, связывающиеся с теми поверхностными белками клетки, к которым прикрепляются ретровирусы, чтобы проникнуть в клетку. Если к такому поверхностному белку уже прикрепился белок ENVV1 или ENVV2, дикий ретровирус не может использовать его для проникновения в клетку.
Таким образом, как минимум три полезных применения нашли себе вирусные гены в плаценте приматов. Это показывает, что генетические модификации, которым ретровирусы подвергают организмы, в долгосрочной перспективе могут оказаться полезными или даже определить развитие вида. И с учетом всего вышесказанного древо доменов должно выглядеть как на схеме ниже (рис. 7).
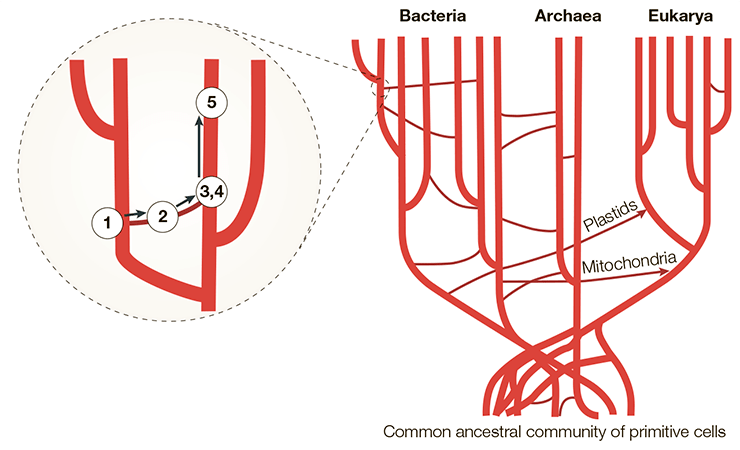
Рисунок 7. Горизонтальный перенос генов в рамках трехдоменного дерева
Заключение
Возникновение паразитов — обязательная черта эволюционирующих систем репликаторов, а соревнование хозяев и паразитов движет эволюцию тех и других. Любой организм является результатом миллионов лет борьбы клеток с невероятно разнообразным миром вирусов. Их действия и их эволюция пронизывают всю историю клеточной эволюции, и сейчас меняется само наше представление о них. Когда-то вирусы считали деградировавшими клетками, но чем больше мы узнаем о вирусах, тем очевиднее, что их роль в общей эволюции значительна. И невероятно много нам еще предстоит узнать.
Статья написана в соавторстве с Евгенией Щепенок.
- Barth F. Smets, Tamar Barkay. (2005). Horizontal gene transfer: perspectives at a crossroads of scientific disciplines. Nat Rev Microbiol. 3, 675-678;
- РНК у истоков жизни?;
- Никитин М.А. (2013). Мир вирусов, последний общий предок и происхождение ДНК. «Химия и жизнь». 8;
- Карл Вёзе (1928-2012);
- Гигантские вирусы: 4-й домен жизни?;
- Элементы: «Формальные статистические тесты подтверждают происхождение всех живых организмов от единого предка»;
- Douglas L. Theobald. (2010). A formal test of the theory of universal common ancestry. Nature. 465, 219-222;
- Никитин М.А. (2013). Происхождение мембран и мембранной биоэнергетики. «Химия и жизнь». 9;
- Кунин Е.В. Логика случая. О прир?